ROCK1
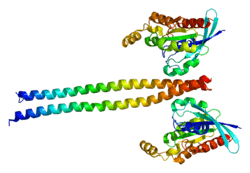
| View/Edit Human | View/Edit Mouse |
ROCK1 is a protein serine/threonine kinase also known as rho-associated, coiled-coil-containing protein kinase 1. Other common names are ROKβ and P160ROCK. ROCK1 is a major downstream effecter of the small GTPase RhoA and is a regulator of the actomyosin cytoskeleton which promotes contractile force generation.[4] ROCK1 plays a role in cancer and in particular cell motility, metastasis, and angiogenesis.[4]
Gene and expression
ROCK1 is also the name of the gene that encodes the protein ROCK1, a serine/threonine kinase. ROCK1 is activated when bound to the GTP-bound form of RhoA. The human ROCK1 gene is located on human chromosome 18 with specific location of 18q11.1.[5] The location of the base pair starts at 18529703 and ends at 18691812 bp and translates into 1354 amino acids.[6]
ROCK1 has a ubiquitous tissue distribution, but subcellularly it is thought to colocalize with the centrosomes. This is consistent with its function as a key modulator of cell motility, tumor cell invasion, and actin cytoskeleton organization.[6] In rats, ROCK1 is expressed in the lung, liver, spleen, kidney, and testis.[7][8][9]
Structure and regulation
The ROCK1 structure is a serine/threonine kinase with molecular weight of 158kDa.[6] It is a homodimer composed of a catalytic kinase domain (residues76-338)[10] located at the amino or N-terminus of the protein, a coiled-coil region(residues 425-1100)[10] containing the Rho-binding domain, and a pleckstrin-homology domain(residues 1118-1317)[10] with a cysteine-rich domain. When a substrate is absent, ROCK1 is an autoinhibited loop structure. Enzyme activity of ROCK1 is inhibited when the pleckstrin-homology and Rho-binding domains in the C-terminus independently bind to the N-terminus kinase domain. When a substrate such as GTP-bound RhoA binds to the Rho-binding region of the coiled-coil domain, the interactions between the N-terminus and the C-terminus are disrupted, thus activating the protein. Cleavage of the C-terminal inhibitory domain by caspase-3 during apoptosis can also activate the kinase.[11]
This view of autoinhibition released by RhoA binding has been challenged by low resolution electron microscopy data showing ROCK to be a constitutive linear dimer 120 nm in length.[12] According to this new data ROCK does not need to be activated by RhoA or phosphorylation because it is always active, and whether ROCK will phosphorylate its substrates (e.g. myosin regulatory light chain) depends only on their subcellular localization.[12]
There is one other isoform of ROCK known as ROCK2. ROCK2 is located at 2p24 and is highly homologous with ROCK1 with an overall amino acid sequence identity of 65%.[10] The identity in the Rho-binding domain is 58%[10] and approximately 92%[10] in the kinase domain. The ROCK isoforms are encoded by two different identified genes and are ubiquitously expressed.[10]
GTPase-RhoA binding can increase the activity of ROCK1 by 1.5-2-fold.[13] Without RhoA binding, lipids such as arachidonic acid or sphingosine phosphorylcholine can increase ROCK1 activity 5- to 6-fold.[13][14] These two lipids interact with the pleckstrin-homology domain, thus disrupting its ability to inhibit ROCK1.[15] G-protein RhoE binds to the N-terminus of ROCK1 and inhibits its activity by preventing RhoA binding. Small G-proteins, Gem and Rad, have been shown to bind and inhibit ROCK1 function, but their mechanism of action is unclear.[10]
Substrates and interactions
ROCK1 phosphorylation sites are at RXXS/T or RXS/T.[10] More than 15 ROCK1 substrates have been identified and activation from these substrates most often leads to actin filament formation and cytoskeleton rearrangements.[10] MYPT-1 is involved in a pathway for smooth muscle contraction. When ROCK1 is activated by binding of GTPase RhoA it produces multiple signaling cascades. For example, RhoA is one of the downstream signaling cascades activated by vascular endothelial growth factor (VEGF). ROCK1 acts as a negative regulator of VEGF endothelial cell activation and angiogensis.[16] ROCK1 activation by RhoA also promotes stabilization of F-actin, phosphorylation of regulatory myosin light chain(MLC) and an increase in contractility, which plays a crucial role in tumor cell migration and metastasis.[17] This activated ROCK1 also activates LIM kinase, which, phosphorylates cofilin, inhibiting its actin-depolymerizing activity.[18] This depolymerization results in stabilization of actin filaments and decreased branching which promotes contraction.
Cardiac troponin is another ROCK1 substrate that upon phosphrylation causes reduction in tension in cardiac myocytes.[10] ROCK1 also acts as a suppressor of inflammatory cell migration by regulating PTEN phosphorylation and stability.
Function
ROCK1 has a diverse range of functions in the body. It is a key regulator of actin-myosin contraction, stability, and cell polarity.[16] These contribute to many progresses such as regulation of morphology, gene transcription, proliferation, differentiation, apoptosis and oncogenic transformation.[4] Other functions involve smooth muscle contraction, actin cytoskeleton organization, stress fiber and focal adhesion formation, neurite retraction, cell adhesion and motility. These functions are activated by phosphorylation of DAPK3, GFAP, LIMK1, LIMK2, MYL9/MLC2, PFN1 and PPP1R12A.[16] Additionally, ROCK1 phosphorylates FHOD1 and acts synergistically with it to promote SRC-dependent non-apoptotic plasma membrane blebbing.[16] It is also required for centrosome positioning and centrosome-dependent exit from mitosis.[16]
Interactions
ROCK1 has been shown to interact with:
Clinical significance
In humans, the main function of ROCK1 is actomyosin contractility. As mentioned before, this contributes to many proximal progresses such as regulation of morphology, motility, and cell–cell and cell–matrix adhesion.[4] In addition, ROCK kinases influence more distal cellular processes including gene transcription, proliferation, differentiation, apoptosis and oncogenic transformation.[4] Given this diverse range of functions is not surprising that ROCK1 has been implicated in numerous aspects of cancer.[4]
Role in cancer
Recent studies have explored the role of ROCK1 in cancer with particular attention focused on cell motility, metastasis, and angiogenesis.[4] Rho GTPases such as RhoA are highly involved in morphologic changes in cells. When a tumor progresses from invasive to metastatic form it requires that they undergo these dramatic morphologic changes. Therefore, increased expression of RhoA and its downstream effector ROCK1 is often observed in human cancers. These cancers are typically more invasive and metastatic phenotypes.[23]
Angiogenesis
Increased expression of RhoA and ROCK1 in endothelial cell migration pathways can cause an increase in angiogenisis and metastatic behavior in tumor cells.[23] It has been suggested that ROCK1 either regulates the expression of angiogenic factors or ROCK1 activation facilitates angiogenesis by increasing the plasticity of the tumor. By reducing the strength of cell-cell interactions and aiding the movement of tumor cells, ROCK1 may enable endothelial cells to penetrate the tumor mass more easily.[23]
Breast cancer
Overexpression of ROCK1 and RhoA is often seen in breast cancer.[24] Activated ROCK1 phosphorylates MLC involved in actin-myosin contractility.[24] RhoA also activates focal adhesion kinase activity. Together, these two pathways create the motile and invasive phenotype of cancer cells. Breast cancers often contain regions of reduced O2 which increases the activity of hypoxia-inducible factors (HIFs). HIFs have been shown to activate transcription of RhoA and ROCK1 leading to cytoskeletoal changes that underlie the invasive cancer cell phenotype.[24]
ROCK1 Inhibitors in cancer therapy
ROCK1 inhibitors might be used in cancer therapy for:
- targeting of stromal rather than tumor cells[10]
- concomitant blocking of ROCK and proteasome activity in K‐Ras‐driven lung cancers [10]
- treating haematological malignancies such as chronic myeloid leukaemia(CML)[10]
ROCK1 inhibition for cancer treatment has not been approved for standard therapy use. Y27632 and Fasudil are examples of ROCK1 inhibitors. Both inhibit ROCK1 by competing with ATP for the kinase activation site. Experiments with Y26732 show it is a promising candidate as a therapeutic antihypertensive agent.[10] Fasudil has been used to characterize the role of ROCK1 in vascular function in clinical studies and has been approved for use in Japan for treatment of cerebral vasospasm following subarachnoid hemorrhage.[10]
Other diseases
The ROCK1 signaling plays an important role in many diseases including diabetes, neurodegenerative diseases such as Parkinson´s disease and amyotrophic lateral sclerosis,[25] and pulmonary hypertension.[26]
See also
References
- ↑ "Drugs that physically interact with Rho-associated protein kinase 1 view/edit references on wikidata".
- ↑ "Human PubMed Reference:".
- ↑ "Mouse PubMed Reference:".
- 1 2 3 4 5 6 7 8 9 10 Rath N, Olson MF (October 2012). "Rho-associated kinases in tumorigenesis: re-considering ROCK inhibition for cancer therapy". EMBO Reports. 13 (10): 900–8. doi:10.1038/embor.2012.127. PMC 3463970
 . PMID 22964758.
. PMID 22964758. - ↑ "ROCK1 Rho-associated, coiled-coil containing protein kinase 1 [ Homo sapiens (human) ]".
- 1 2 3 "Rho-Associated, Coiled-Coil Containing Protein Kinase 1".
- ↑ Hahmann C, Schroeter T (January 2010). "Rho-kinase inhibitors as therapeutics: from pan inhibition to isoform selectivity". Cellular and Molecular Life Sciences. 67 (2): 171–7. doi:10.1007/s00018-009-0189-x. PMID 19907920.
- ↑ Riento K, Ridley AJ (June 2003). "Rocks: multifunctional kinases in cell behaviour". Nature Reviews. Molecular Cell Biology. 4 (6): 446–56. doi:10.1038/nrm1128. PMID 12778124.
- ↑ Nakagawa O, Fujisawa K, Ishizaki T, Saito Y, Nakao K, Narumiya S (August 1996). "ROCK-I and ROCK-II, two isoforms of Rho-associated coiled-coil forming protein serine/threonine kinase in mice". FEBS Letters. 392 (2): 189–93. doi:10.1016/0014-5793(96)00811-3. PMID 8772201.
- 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 "Chapter 7Rho Kinase in Vascular Smooth Muscle".
- ↑ Jacobs M, Hayakawa K, Swenson L, Bellon S, Fleming M, Taslimi P, Doran J (January 2006). "The structure of dimeric ROCK I reveals the mechanism for ligand selectivity". The Journal of Biological Chemistry. 281 (1): 260–8. doi:10.1074/jbc.M508847200. PMID 16249185.
- 1 2 Truebestein L, Elsner DJ, Fuchs E, Leonard TA (2015-12-01). "A molecular ruler regulates cytoskeletal remodelling by the Rho kinases". Nature Communications. 6: 10029. doi:10.1038/ncomms10029. PMID 26620183.
- 1 2 Feng J, Ito M, Kureishi Y, Ichikawa K, Amano M, Isaka N, Okawa K, Iwamatsu A, Kaibuchi K, Hartshorne DJ, Nakano T (February 1999). "Rho-associated kinase of chicken gizzard smooth muscle". The Journal of Biological Chemistry. 274 (6): 3744–52. doi:10.1074/jbc.274.6.3744. PMID 9920927.
- ↑ Shirao S, Kashiwagi S, Sato M, Miwa S, Nakao F, Kurokawa T, Todoroki-Ikeda N, Mogami K, Mizukami Y, Kuriyama S, Haze K, Suzuki M, Kobayashi S (July 2002). "Sphingosylphosphorylcholine is a novel messenger for Rho-kinase-mediated Ca2+ sensitization in the bovine cerebral artery: unimportant role for protein kinase C". Circulation Research. 91 (2): 112–9. doi:10.1161/01.res.0000026057.13161.42. PMID 12142343.
- ↑ Amano M, Fukata Y, Kaibuchi K (November 2000). "Regulation and functions of Rho-associated kinase". Experimental Cell Research. 261 (1): 44–51. doi:10.1006/excr.2000.5046. PMID 11082274.
- 1 2 3 4 5 "Q13464 (ROCK1_HUMAN)".
- ↑ "Inhibition of Rho-dependent kinases ROCK I/II activates VEGF-driven retinal neovascularization and sprouting angiogenesis".
- ↑ "Entrez Gene: ROCK1 Rho-associated, coiled-coil containing protein kinase 1".
- ↑ Da Silva JS, Medina M, Zuliani C, Di Nardo A, Witke W, Dotti CG (September 2003). "RhoA/ROCK regulation of neuritogenesis via profilin IIa-mediated control of actin stability". The Journal of Cell Biology. 162 (7): 1267–79. doi:10.1083/jcb.200304021. PMC 2173969. PMID 14517206.
- ↑ Riento K, Guasch RM, Garg R, Jin B, Ridley AJ (June 2003). "RhoE binds to ROCK I and inhibits downstream signaling". Molecular and Cellular Biology. 23 (12): 4219–29. doi:10.1128/mcb.23.12.4219-4229.2003. PMC 156133. PMID 12773565.
- ↑ Leung T, Chen XQ, Manser E, Lim L (October 1996). "The p160 RhoA-binding kinase ROK alpha is a member of a kinase family and is involved in the reorganization of the cytoskeleton". Molecular and Cellular Biology. 16 (10): 5313–27. PMC 231530. PMID 8816443.
- ↑ Fujisawa K, Fujita A, Ishizaki T, Saito Y, Narumiya S (September 1996). "Identification of the Rho-binding domain of p160ROCK, a Rho-associated coiled-coil containing protein kinase". The Journal of Biological Chemistry. 271 (38): 23022–8. doi:10.1074/jbc.271.38.23022. PMID 8798490.
- 1 2 3 Croft DR, Sahai E, Mavria G, Li S, Tsai J, Lee WM, Marshall CJ, Olson MF (December 2004). "Conditional ROCK activation in vivo induces tumor cell dissemination and angiogenesis". Cancer Research. 64 (24): 8994–9001. doi:10.1158/0008-5472.CAN-04-2052. PMID 15604264.
- 1 2 3 Gilkes DM, Xiang L, Lee SJ, Chaturvedi P, Hubbi ME, Wirtz D, Semenza GL (January 2014). "Hypoxia-inducible factors mediate coordinated RhoA-ROCK1 expression and signaling in breast cancer cells". Proceedings of the National Academy of Sciences of the United States of America. 111 (3): E384–93. doi:10.1073/pnas.1321510111. PMC 3903228. PMID 24324133.
- ↑ Tönges L, Frank T, Tatenhorst L, Saal KA, Koch JC, Szego É, Bähr M, Weishaupt JH, Lingor P (November 2012). "Inhibition of rho kinase enhances survival of dopaminergic neurons and attenuates axonal loss in a mouse model of Parkinson's disease". Brain. 135 (Pt 11): 3355–70. doi:10.1093/brain/aws254. PMC 3501973. PMID 23087045.
- ↑ Dahal BK, Kosanovic D, Pamarthi PK, Sydykov A, Lai YJ, Kast R, Schirok H, Stasch JP, Ghofrani HA, Weissmann N, Grimminger F, Seeger W, Schermuly RT (October 2010). "Therapeutic efficacy of azaindole-1 in experimental pulmonary hypertension". The European Respiratory Journal. 36 (4): 808–18. doi:10.1183/09031936.00140309. PMID 20530035.
Further reading
- Riento K, Ridley AJ (June 2003). "Rocks: multifunctional kinases in cell behaviour". Nature Reviews. Molecular Cell Biology. 4 (6): 446–56. doi:10.1038/nrm1128. PMID 12778124.
External links
- ROCK1 Info with links in the Cell Migration Gateway
PDB gallery | |
|---|---|