Bat
| Bat Temporal range: Eocene – Present | |
|---|---|
 | |
| Clockwise: common Egyptian fruit bat Rousettus aegyptiacus, Mexican free-tail bat Tadarida brasiliensis, Myotis myotis, lesser short-nose fruit bat Cynopterus sphinx, horseshoe bat Rhinolophus ferrumequinum, common vampire bat Desmodus rotundus. | |
| Scientific classification | |
| Kingdom: | Animalia |
| Phylum: | Chordata |
| Class: | Mammalia |
| Clade: | Scrotifera |
| Order: | Chiroptera Blumenbach, 1779 |
| Suborders | |
 | |
| Worldwide distribution of bat species | |


Bats are mammals of the order Chiroptera (/kaɪˈrɒptərə/; from the Ancient Greek: χείρ - cheir, "hand"[1] and Ancient Greek: πτερόν - pteron, "wing"[2])[3] whose forelimbs form webbed wings, making them the only mammals naturally capable of true and sustained flight.[3] By contrast, other mammals said to fly, such as flying squirrels, gliding possums, and colugos, can only glide for short distances. Bats do not flap their entire forelimbs, as birds do, but instead flap their spread-out digits,[4] which are very long and covered with a thin membrane or patagium.
Bats are the second largest order of mammals (after the rodents), representing about 20% of all classified mammal species worldwide, with about 1,240 bat species divided into two suborders: the less specialized and largely fruit-eating megabats, or flying foxes, and the highly specialized and echolocating microbats.[5] About 70% of bat species are insectivores. Most of the rest are frugivores, or fruit eaters. A few species, such as the fish-eating bat, feed from animals other than insects, with the vampire bats being hematophagous, or feeding on blood.
Bats are present throughout most of the world, with the exception of extremely cold regions. They perform the vital ecological roles of pollinating flowers and dispersing fruit seeds; many tropical plant species depend entirely on bats for the distribution of their seeds. Bats are economically important, as they consume insect pests, reducing the need for pesticides. The smallest bat is the Kitti's hog-nosed bat, measuring 29–34 mm (1.14–1.34 in) in length, 15 cm (5.91 in) across the wings and 2–2.6 g (0.07–0.09 oz) in mass.[6][7] It is also arguably the smallest extant species of mammal, with the Etruscan shrew being the other contender.[8] The largest species of bat are a few species of Pteropus (fruit bats or flying foxes) and the giant golden-crowned flying fox with a weight up to 1.6 kg (4 lb) and wingspan up to 1.7 m (5 ft 7 in).[9]
The Mexican free-tailed bat is the fastest flying animal in horizontal flight.[10]
Etymology
In many languages, the word for "bat" is cognate with the word for "mouse": for example, chauve-souris ("bald-mouse") in French, murciélago ("blind mouse") in Spanish, saguzahar ("old mouse") in Basque, летучая мышь ("flying mouse") in Russian, slijepi miš ("blind mouse") in Bosnian, nahkhiir ("leather mouse") in Estonian, vlermuis (winged mouse) in Afrikaans, from the Dutch word vleermuis (from Middle Dutch "winged mouse").
An older English name for bats is flittermouse, which matches their name in other Germanic languages (for example German Fledermaus and Swedish fladdermus), related to fluttering of wings. Middle English had bakke, which may have undergone a shift from -k- to -t- influenced by Latin blatta, "moth, nocturnal insect".[11]
Classification and evolution

Bats are placental mammals. Bats were formerly thought to have been most closely related to the flying lemurs, treeshrews, and primates,[12] but recent molecular cladistics research indicates that they actually belong to Laurasiatheria, a diverse group also containing Carnivora and Artiodactyla.[13][14]
The two traditionally recognized suborders of bats are:
- Megachiroptera (megabats)
- Microchiroptera (microbats/echolocating bats)
Not all megabats are larger than microbats. The major distinctions between the two suborders are:
- Microbats use echolocation; with the exception of the genus Rousettus, megabats do not.[15]
- Microbats lack the claw at the second finger of the forelimb.[16][17]
- The ears of microbats do not close to form a ring; the edges are separated from each other at the base of the ear.[17]
- Microbats lack underfur; they are either naked or have guard hairs.
Megabats eat fruit, nectar, or pollen. Most microbats eat insects; others may feed on fruit, nectar, pollen, fish, frogs, small mammals, or the blood of animals. Megabats have well-developed visual cortices and show good visual acuity, while microbats rely on echolocation for navigation and finding prey.
The phylogenetic relationships of the different groups of bats have been the subject of much debate. The traditional subdivision between Megachiroptera and Microchiroptera reflects the view that these groups of bats have evolved independently of each other for a long time, from a common ancestor already capable of flight. This hypothesis recognized differences between microbats and megabats and acknowledged that flight has only evolved once in mammals. Most molecular biological evidence supports the view that bats form a single or monophyletic group.[18]
Researchers have proposed alternative views of chiropteran phylogeny and classification, but more research is needed.
In the 1980s, a hypothesis based on morphological evidence was offered that stated the Megachiroptera evolved flight separately from the Microchiroptera. The so-called flying primate hypothesis proposes that, when adaptations to flight are removed, the Megachiroptera are allied to primates by anatomical features not shared with Microchiroptera. One example is that the brains of megabats show a number of advanced characteristics that link them to primates. Although recent genetic studies strongly support the monophyly of bats,[19] debate continues as to the meaning of available genetic and morphological evidence.[20]
Genetic evidence indicates that megabats originated during the early Eocene and should be placed within the four major lines of microbats.[21]
Consequently, two new suborders based on molecular data have been proposed. The new suborder of Yinpterochiroptera includes the Pteropodidae, or megabat family, as well as the families Rhinolophidae, Hipposideridae, Craseonycteridae, Megadermatidae, and Rhinopomatidae[22] The other new suborder, Yangochiroptera, includes all of the remaining families of bats (all of which use laryngeal echolocation). These two new suborders are strongly supported by statistical tests. Teeling (2005) found 100% bootstrap support in all maximum likelihood analyses for the division of Chiroptera into these two modified suborders. This conclusion is further supported by a 15-base-pair deletion in BRCA1 and a seven-base-pair deletion in PLCB4 present in all Yangochiroptera and absent in all Yinpterochiroptera.[22] Perhaps most convincingly, a phylogenomic study by Tsagkogeorga et al (2013) showed that the two new proposed suborders were supported by analyses of thousands of genes.[21]
The chiropteran phylogeny based on molecular evidence is controversial because microbat paraphyly implies that one of two seemingly unlikely hypotheses occurred. The first suggests that laryngeal echolocation evolved twice in Chiroptera, once in Yangochiroptera and once in the rhinolophoids.[23][24] The second proposes that laryngeal echolocation had a single origin in Chiroptera, was subsequently lost in the family Pteropodidae (all megabats), and later evolved as a system of tongue-clicking in the genus Rousettus.[25]

Analyses of the sequence of the "vocalization" gene, FoxP2, were inconclusive as to whether laryngeal echolocation was secondarily lost in the pteropodids or independently gained in the echolocating lineages.[26] However, analyses of the "hearing" gene, Prestin, seemed to favor the independent gain in echolocating species rather than a secondary loss in the pteropodids.[27]
In addition to Yinpterochiroptera and Yangochiroptera, the names Pteropodiformes and Vespertilioniformes have also been proposed for these suborders.[28][29] Under this new proposed nomenclature, the suborder Pteropodiformes includes all extant bat families more closely related to the genus Pteropus than the genus Vespertilio, while the suborder Vespertilioniformes includes all extant bat families more closely related to the genus Vespertilio than to the genus Pteropus.
Little fossil evidence is available to help map the evolution of bats, since their small, delicate skeletons do not fossilize very well. However, a Late Cretaceous tooth from South America resembles that of an early microchiropteran bat. Most of the oldest known, definitely identified bat fossils were already very similar to modern microbats. These fossils, Icaronycteris, Archaeonycteris, Palaeochiropteryx and Hassianycteris, are from the early Eocene period, 52.5 million years ago.[18] Archaeopteropus, formerly classified as the earliest known megachiropteran, is now classified as a microchiropteran.
Bats were formerly grouped in the superorder Archonta, along with the treeshrews (Scandentia), colugos (Dermoptera), and the primates, because of the apparent similarities between Megachiroptera and such mammals. Genetic studies have now placed bats in the superorder Laurasiatheria, along with carnivorans, pangolins, odd-toed ungulates, even-toed ungulates, and cetaceans.[30] A recent study by Zhang et al. places Chiroptera as a sister taxon to the clade Perissodactyla (which includes horses and other odd-toed ungulates).[31] However, the first phylogenomic analysis of bats shows that they are not sisters to Perissodactyla, instead they are sisters to a larger group that includes ungulates and carnivores.[21]
| Boreoeutheria |
| ||||||||||||||||||||||||||||||||||||
| |

The traditional classification of bats is:
- Order Chiroptera
- Suborder Megachiroptera (megabats)
- Suborder Microchiroptera (microbats)
- Superfamily Emballonuroidea
- Emballonuridae (Sac-winged or sheath-tailed bats)
- Superfamily Molossoidea
- Superfamily Nataloidea
- Superfamily Noctilionoidea
- Mormoopidae (Ghost-faced or moustached bats)
- Mystacinidae (New Zealand short-tailed bats)
- Noctilionidae (Bulldog bats or fisherman bats)
- Phyllostomidae (Leaf-nosed bats)
- Superfamily Rhinolophoidea
- Megadermatidae (False vampires)
- Nycteridae (Hollow-faced or slit-faced bats)
- Rhinolophidae (Horseshoe bats)
- Hipposideridae (Old World leaf-nosed bats)
- Superfamily Rhinopomatoidea
- Craseonycteridae (Bumblebee bat or Kitti's hog-nosed bat)
- Rhinopomatidae (Mouse-tailed bats)
- Superfamily Vespertilionoidea
- Vespertilionidae (Vesper bats or evening bats)
- Antrozoidae (Pallid bat and Van Gelder's bat)
- Superfamily Emballonuroidea
Megabats primarily eat fruit or nectar. In New Guinea, they are likely to have evolved for some time in the absence of microbats, which has resulted in some smaller megabats of the genus Nyctimene becoming (partly) insectivorous to fill the vacant microbat ecological niche. Furthermore, some evidence indicates that the fruit bat genus Pteralopex from the Solomon Islands, and its close relative Mirimiri from Fiji, have evolved to fill some niches that were open because there are no nonvolant or nonflying mammals on those islands.
Fossil bats
Fossilized remains of bats are few, as they are terrestrial and light-boned. Only an estimated 12% of the bat fossil record is complete at the genus level.[32] Fossil remains of an Eocene bat, Icaronycteris, were found in 1960. Another Eocene bat, Onychonycteris finneyi, was found in the 52-million-year-old Green River Formation in Wyoming, United States, in 2003.[33][34] This intermediate fossil has helped to resolve a long-standing disagreement regarding whether flight or echolocation developed first in bats. The shape of the rib cage, faceted infraspious fossa of the scapula, manus morphology, robust clavicle, and keeled sternum all indicated Onychonycteris was capable of powered flight. However, the well-preserved skeleton showed that the small cochlea of the inner ear did not have the morphology necessary to echolocate. O. finneyi lacked an enlarged orbical apophysis on the malleus, and a stylohyal element with an expanded paddle-like cranial tip—both of which are characteristics linked to echolocation in other prehistoric and extant bat species.[18] Because of these absences, and the presence of characteristics necessary for flight, Onychonycteris provides strong support for the “flight first” hypothesis in the evolution of flight and echolocation in bats.
The appearance and flight movement of bats 52.5 million years ago were different from those of bats today. Onychonycteris had claws on all five of its fingers, whereas modern bats have at most two claws appearing on two digits of each hand. It also had longer hind legs and shorter forearms, similar to climbing mammals that hang under branches, such as sloths and gibbons. This palm-sized bat had short, broad wings, suggesting that it could not fly as fast or as far as later bat species. Instead of flapping its wings continuously while flying, Onychonycteris likely alternated between flaps and glides while in the air.[18] Such physical characteristics suggest that this bat did not fly as much as modern bats do, rather flying from tree to tree and spending most of its waking day climbing or hanging on the branches of trees.[35] The distinctive features noted on the Onychonycteris fossil also support the claim that mammalian flight most likely evolved in arboreal gliders, rather than terrestrial runners. This model of flight development, commonly known as the "trees-down" theory, implies that bats attained powered flight by taking advantage of height and gravity, rather than relying on running speeds fast enough for a ground-level take off.[36]
The mid-Eocene genus Necromantis is one of the earliest examples of bats specialised to hunt vertebrate prey, as well as one of the largest bats of its epoch. The late-Eocene Witwatia is another similarly large predatory bat, while Aegyptonycteris is among the first and largest omnivorous bat species.
The extinct bats Palaeochiropteryx tupaiodon and Hassianycteris kumari are the first fossil mammals to have their colouration discovered, both of a reddish brown.[37]
Habitats
Flight has enabled bats to become one of the most widely distributed groups of mammals.[38] Apart from the Arctic, the Antarctic and a few isolated oceanic islands, bats exist all over the world.[39] Bats are found in almost every habitat available on Earth. Different species select different habitats during different seasons, ranging from seasides to mountains and even deserts, but bat habitats have two basic requirements: roosts, where they spend the day or hibernate, and places for foraging. Most temperate species additionally need a relatively warm hibernation shelter. Bat roosts can be found in hollows, crevices, foliage, and even human-made structures, and include "tents" the bats construct by biting leaves.[40]
The United States is home to an estimated 45 to 48 species of bats.[41][42] The three most common species are Myotis lucifugus (little brown bat), Eptesicus fuscus (big brown bat), and Tadarida brasiliensis (Mexican free-tailed bat). The little and the big brown bats are common throughout the northern two-thirds of the country, while the Mexican free-tailed bat is the most common species in the southwest,[43] sometimes even appearing in portions of the Southeast.
Anatomy
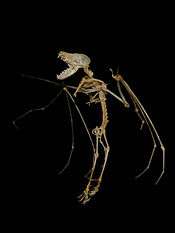
Wings


The finger bones of bats are much more flexible than those of other mammals, owing to their flattened cross-section and to low levels of minerals, such as calcium, near their tips. In 2006, Sears et al. published a study that traces the elongation of manual bat digits, a key feature required for wing development, to the upregulation of bone morphogenetic proteins (Bmps). During embryonic development, the gene controlling Bmp signaling, Bmp2, is subjected to increased expression in bat forelimbs—resulting in the extension of the offspring's manual digits. This crucial genetic alteration helps create the specialized limbs required for volant locomotion.[44] Sears et al. (2006) also studied the relative proportion of bat forelimb digits from several extant species and compared these with a fossil of Lcaronycteris index, an early extinct species from approximately 50 million years ago. The study found no significant differences in relative digit proportion, suggesting that bat wing morphology has been conserved for over 50 million years.[44]
The wings of bats are much thinner and consist of more bones than the wings of birds, allowing bats to maneuver more accurately than the latter, and fly with more lift and less drag.[45] By folding the wings in toward their bodies on the upstroke, they save 35 percent energy during flight.[46] The membranes are also delicate, ripping easily;[47] however, the tissue of the bat's membrane is able to regrow, such that small tears can heal quickly.[47][48] The surface of their wings is equipped with touch-sensitive receptors on small bumps called Merkel cells, also found on human fingertips. These sensitive areas are different in bats, as each bump has a tiny hair in the center,[49] making it even more sensitive and allowing the bat to detect and collect information about the air flowing over its wings, and to fly more efficiently by changing the shape of its wings in response.[49] An additional kind of receptor cell is found in the wing membrane of species that use their wings to catch prey. This receptor cell is sensitive to the stretching of the membrane.[49] The cells are concentrated in areas of the membrane where insects hit the wings when the bats capture them.
Circulatory system
Bats seem to make use of particularly strong venomotion (rhythmic contraction of venous wall muscles). In most mammals, the walls of the veins provide mainly passive resistance (maintaining their shape as deoxygenated blood flows through them), but in bats they appear to actively support blood flow back to the heart with this pumping action.[50][51]
Bats also possess a system of sphincter valves on the arterial side of the vascular network that runs along the edge of their wings. In the fully open state, these allow oxygenated blood to flow through the capillary network across the flight membrane (i.e. wing surface), but when contracted, they shunt flow directly to the veins, bypassing the wing capillaries. This is likely an important tool for thermoregulation, allowing the bats to control the amount of heat exchanged through the thin flight membrane (many other mammals use the capillary network in oversized ears for the same purpose).[52]
Other
The teeth of microbats resemble insectivorans. They are very sharp to bite through the hardened armor of insects or the skin of fruit.
The tube-lipped nectar bat (Anoura fistulata) has the longest tongue of any mammal relative to its body size. This is beneficial to them in terms of pollination and feeding. Their long, narrow tongues can reach deep into the long cup shape of some flowers. When the tongue retracts, it coils up inside its rib cage.[53]
Bats possess highly adapted lung systems to cope with the pressures of powered-flight. Flight is an energetically taxing aerobic activity and requires large amounts of oxygen to be sustained. In bats, the relative alveolar surface area and pulmonary capillary blood volume are significantly larger than most other small quadrupedal mammals.[54]
Echolocation
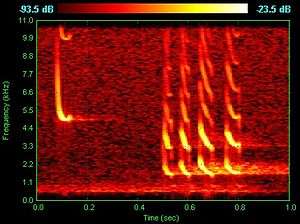
 |
Pipistrellus Pulses
 Recording of Pipistrellus pipistrellus bat time-expanded echolocation calls and social call. |
| Problems playing this file? See media help. | |
Bat echolocation is a perceptual system where ultrasonic sounds are emitted specifically to produce echoes. By comparing the outgoing pulse with the returning echoes, the brain and auditory nervous system can produce detailed images of the bat's surroundings. This allows bats to detect, localize, and even classify their prey in complete darkness. At 130 decibels in intensity, bat calls are some of the most intense, airborne animal sounds.[56]
To clearly distinguish returning information, bats must be able to separate their calls from the echoes that they receive. Microbats use two distinct approaches.
- Low duty cycle echolocation: Bats can separate their calls and returning echoes by time. Bats that use this approach time their short calls to finish before echoes return. This is important because these bats contract their middle ear muscles when emitting a call, so they can avoid deafening themselves. The time interval between the call and echo allows them to relax these muscles, so they can clearly hear the returning echo.[57] The delay of the returning echoes provides the bat with the ability to estimate the range to their prey.
- High duty cycle echolocation: Bats emit a continuous call and separate pulse and echo in frequency. The ears of these bats are sharply tuned to a specific frequency range. They emit calls outside of this range to avoid self-deafening. They then receive echoes back at the finely tuned frequency range by taking advantage of the Doppler shift of their motion in flight. The Doppler shift of the returning echoes yields information relating to the motion and location of the bat's prey. These bats must deal with changes in the Doppler shift due to changes in their flight speed. They have adapted to change their pulse emission frequency in relation to their flight speed so echoes still return in the optimal hearing range.[58]
The new Yinpterochiroptera and Yangochiroptera classification of bats, supported by molecular evidence, suggests two possibilities for the evolution of echolocation. It may have been gained once in a common ancestor of all bats and was then subsequently lost in the Old World fruit bats, only to be regained in the horseshoe bats, or echolocation evolved independently in both the Yinpterochiroptera and Yangochiroptera lineages.[59]
Two groups of moths exploit a bat sense to echolocate: tiger moths produce ultrasonic signals to warn the bats that they (the moths) are chemically protected or aposematic, other moth species produce signals to jam bat echolocation. Many moth species have a hearing organ called a tympanum, which responds to an incoming bat signal by causing the moth's flight muscles to twitch erratically, sending the moth into random evasive maneuvers.

In addition to echolocating prey, bat ears are sensitive to the fluttering of moth wings, the sounds produced by tymbalate insects, and the movement of ground-dwelling prey, such as centipedes, earwigs, etc. The complex geometry of ridges on the inner surface of bat ears helps to sharply focus not only echolocation signals, but also to passively listen for any other sound produced by the prey. These ridges can be regarded as the acoustic equivalent of a Fresnel lens, and may be seen in a large variety of unrelated animals, such as the aye-aye, lesser galago, bat-eared fox, mouse lemur, and others.[60][61][62]
By repeated scanning, bats can mentally construct an accurate image of the environment in which they are moving and of their prey item.[63]
Other senses
Although the eyes of most microbat species are small and poorly developed, leading to poor visual acuity, no species is blind.[64] Microbats use vision to navigate, especially for long distances when beyond the range of echolocation,[65] and species that are gleaners—that is, ones that attempt to swoop down from above to ambush insects, like crickets on the ground or moths up a tree, often have eyesight about as good as a rat's. Some species have been shown to be able to detect ultraviolet light, and most cave-dwelling species have developed the ability to utilize very dim light. They also have high-quality senses of smell and hearing. Bats hunt at night, reducing competition with birds, minimizing contact with certain predators, and travel large distances (up to 800 km) in their search for food.[4]
Megabat species often have excellent eyesight as good as, if not better than, human vision. This eyesight is, unlike its microbat relations, adapted to both night and daylight vision and enables the bat to have some colour vision whereas the microbat sees in blurred shades of grey.
Behaviour
Most microbats are nocturnal[66] and are active at twilight. A large portion of bats migrate hundreds of kilometres to winter hibernation dens,[67] while some pass into torpor in cold weather, rousing and feeding when warm weather allows for insects to be active.[68] Others retreat to caves for winter and hibernate for six months.[68] Bats rarely fly in rain, as the rain interferes with their echolocation, and they are unable to locate their food.
The social structure of bats varies, with some leading solitary lives and others living in caves colonized by more than a million bats.[69] The fission-fusion social structure is seen among several species of bats. The term "fusion" refers to a large numbers of bats that congregate in one roosting area, and "fission" refers to breaking up and the mixing of subgroups, with individual bats switching roosts with others and often ending up in different trees and with different roostmates.
Studies also show that bats make various sounds in order to communicate with one another. Scientists in the field have listened to bats and have been able to associate certain sounds with certain behaviours that bats make after the sounds are made.[69]
Insectivores make up 70% of bat species and locate their prey by means of echolocation. Of the remainder, most feed on fruits.[70] Only three species sustain themselves with blood.
Some species even prey on vertebrates. The leaf-nosed bats (Phyllostomidae) of Central America and South America, and the two bulldog bat (Noctilionidae) species feed on fish. At least two species of bat are known to feed on other bats: the spectral bat, also known as the American false vampire bat, and the ghost bat of Australia.[70] One species, the greater noctule bat, catches and eats small birds in the air.
Predators of bats include bat hawks, bat falcons and even spiders.[71]
Reproduction


Most bats have a breeding season, which is in the spring for species living in a temperate climate. Bats may have one to three litters in a season, depending on the species and on environmental conditions, such as the availability of food and roost sites. Females generally have one offspring at a time, which could be a result of the mother's need to fly to feed while pregnant. Female bats nurse their young until they are nearly adult size, because a young bat cannot forage on its own until its wings are fully developed.
Female bats use a variety of strategies to control the timing of pregnancy and the birth of young, to make delivery coincide with maximum food ability and other ecological factors. Females of some species have delayed fertilization, in which sperm is stored in the reproductive tract for several months after mating. In many such cases, mating occurs in the fall, and fertilization does not occur until the following spring. Other species exhibit delayed implantation, in which the egg is fertilized after mating, but remains free in the reproductive tract until external conditions become favorable for giving birth and caring for the offspring.
In yet another strategy, fertilization and implantation both occur, but development of the fetus is delayed until favorable conditions prevail, during the delayed development the mother still gives the fertilized egg nutrients, and oxygenated blood to keep it alive. However, this process can go for a long period of time, because of the advanced gas exchange system.[72] All of these adaptations result in the pup being born during a time of high local production of fruit or insects.
At birth, the wings are too small to be used for flight. Young microbats become independent at the age of six to eight weeks, while megabats do not until they are four months old.
Life expectancy
A single bat can live over 20 years, but bat population growth is limited by the slow birth rate.[73] Five species have been recorded living over 30 years in the wild: the brown long-eared bat (Plecotus auritus), little brown bat (Myotis lucifugus), Brandt's bat (Myotis brandti), lesser mouse-eared bat (Myotis blythii) and greater horseshoe bat (Rhinolophus ferrumequinum).[74]
Hunting, feeding, and drinking

Newborn bats feed solely on their mother's milk.[75] When they are a few weeks old, bats are expected to fly and hunt on their own. It is up to them to find and catch their prey, along with satisfying their thirst.[76]
To survive hibernation months, some species build up large reserves of body fat, both as fuel and as insulation.[76]
Hunting

Most bats are nocturnal creatures. Their daylight hours are spent grooming and sleeping; they hunt during the night. The means by which bats navigate while finding and catching their prey in the dark was unknown until the 1790s, when Lazzaro Spallanzani conducted a series of experiments on a group of hooded and surgically blinded bats.[77] These bats were placed in a room in total darkness, with silk threads strung across the room. Even then, the bats were able to navigate their way through the room. Spallanzani concluded the bats were not using their eyes to fly through complete darkness, but something else.
Spallanzani decided the bats were able to catch and find their prey through the use of their ears. To prove this theory, Spallanzani plugged the ears of the bats in his experiment. To his pleasure, he found that the bats with plugged ears were not able to fly with the same amount of skill and precision as they were able to without their ears plugged. Unfortunately for Spallanzani, the twin concepts of sound waves and acoustics would not be understood for another century and he could not explain why specifically the bats were crashing into walls and the threads that he'd strung up around the room, and because of the methodology Spallanzani used, many of his test subjects died.
It was thus well known through the nineteenth century that the chiropteran ability to navigate had something to do with hearing, but how they accomplish this was not proven conclusively until the 1930s, by Donald R. Griffin, a biology student at Harvard University. Using a locally native species, the little brown bat, he discovered that bats use echolocation to locate and catch their prey. When bats fly, they produce a constant stream of high-pitched sounds. When the sound waves produced by these sounds hit an insect or other animal, the echoes bounce back to the bat, and guide them to the source.[76]
Feeding and diet
The majority of food consumed by bats includes insects, fruits and flower nectar, vertebrates and blood.[78] Almost three-fourths of the world's bats are insect eaters. Bats consume both aerial and ground-dwelling insects. Each bat is typically able to consume one-third of its body weight in insects each night, and several hundred insects in a few hours. This means that a group of a thousand bats could eat four tons of insects each year. If bats were to become extinct, it has been calculated that the insect population would reach an alarmingly high number.[79]
Vitamin C
The Chiroptera as a whole are in the process of losing the ability to synthesize vitamin C: most have lost it completely.[80] In a test of 34 bat species from six major families of bats, including major insect- and fruit-eating bat families, all were found to have lost the ability to synthesize it, and this loss may derive from a common bat ancestor, as a single mutation.[81] However, recent results show that there are at least two species of bat, the frugivorous bat (Rousettus leschenaultii) and insectivorous bat (Hipposideros armiger), that have retained their ability to produce vitamin C.[82]
Aerial insectivores
Watching a bat catch and eat an insect is difficult. The action is so fast that all one sees is a bat rapidly changing directions, and continuing on its way. Scientist Frederick A. Webster discovered how bats catch their prey. In 1960, Webster developed a high-speed camera that was able to take one thousand pictures per second. These photos revealed the fast and precise way in which bats catch insects.[76] Occasionally, a bat will catch an insect in mid-air with its mouth, and eat it in the air. However, more often than not, a bat will use its tail membrane or wings to scoop up the insect and trap it in a sort of "bug net".[75] Then, the bat will take the insect back to its roost. There, the bat will proceed to eat said insect, often using its tail membrane as a kind of napkin, to prevent its meal from falling to the ground.[78] One common insect prey is Helicoverpa zea, a moth that causes major agricultural damage.[83]
Forage gleaners
These bats typically fly down and grasp their prey off the ground with their teeth, and take it to a nearby perch to eat it. Generally, these bats do not use echolocation to locate their prey. Instead, they rely on the sounds produced by the insects. Some make unique sounds, and almost all make some noise while moving through the environment.[75]
Fruits and flower nectar
.jpg)
Fruit eating, or frugivory, is found in particular species from both major suborders. These bats favor fleshy and sweet fruits, but not those particularly strong smelling or colorful.[75] They pull the fruit off the trees with their teeth, then fly back to their roosts to consume them, sucking out the juice and spitting the seeds and pulp out onto the ground.
This helps disperse the seeds of these fruit trees, which may take root and grow where the bats have left them.[79] Over 150 types of plants depend on bats in order to reproduce.[79]
Some Chiropterans consume nectar instead, for which they have acquired specialized adaptations. These bats possess long muzzles and long, extensible tongues covered in fine bristles that aid them in feeding on particular flowers and plants.[75] However, because of these features, nectar-feeding bats cannot easily turn to other food sources in times of scarcity, making them more prone to extinction than any other type of bat.[84][85]
Nectar feeding also aids a variety of plants, since these bats serve as pollinators: pollen gets stuck to the bats' fur while they sip the nectar, and is transferred to the next flower they visit (or dusts off in flight).[79] Rainforests are said to benefit the most from bat pollination, because of the large variety of plants that depend on it.[86]
Vertebrates
Some bats are primarily carnivorous, feeding on vertebrates.[75] These bats typically eat a variety of animals, especially frogs, lizards, birds, and sometimes other bats.[79]
Trachops cirrhosus, for example, is particularly skilled at catching frogs. These bats locate large groups of frogs by tracking their mating calls, then plucking them from the surface of the water with their sharp canine teeth.[75] Another example is the greater noctule bat, which is believed to catch birds in flight.
Also, several bat species, found on all continents, feed on fish. They use echolocation to detect tiny ripples on the water's surface, swoop down and use specially enlarged claws on their hind feet to grab the fish, then take their prey to a feeding roost and consume it.[75]
Blood
A few species, specifically the common, white-winged, and hairy-legged vampire bats, exclusively consume animal blood. This is referred to as hematophagy. The common vampire bat typically feeds on mammals, while the hairy-legged and white-winged vampires feed on birds instead.[87] These species are found throughout Central and South America, as well as in Mexico and on the island of Trinidad.
Defecation
Bat dung, or guano, is so rich in nutrients that it is mined from caves, bagged, and used by farmers to fertilize their crops. During the U.S. Civil War, guano was used to make gunpowder.[79]
Drinking
In 1960, Frederic A. Webster discovered some bats' method of drinking water using a high-speed (1000 FPS) camera and flashgun. He captured one skimming just above the surface of the water, lowering its jaw to collect a small quantity of water on each pass, taking repeated passes until it drank its fill.[76]
Other bats, such as the flying fox or fruit bat, gently skim the water's surface, then land nearby to lick the water from their chest fur.[88]
Interactions with humans
Disease transmission
Bats are natural reservoirs for a large number of zoonotic pathogens,[89] including rabies,[90] histoplasmosis (directly and in guano),[91] Henipavirus (i.e. Nipah virus and Hendra virus)[92] and possibly ebola virus.[93][94]
Their high mobility, broad distribution, long life spans, substantial sympatry, and social behaviour (communal roosting and fission-fusion social structure) make bats favourable hosts and vectors of disease. Compared to rodents, bats carry more zoonotic viruses per species, and each virus is shared with more (especially sympatric) species.[95] They also seem to be highly resistant to many of the pathogens they carry, suggesting a potential commensal/mutualistic relationship or specific adaptations to the bats' immune systems.[95][96][97] Furthermore, their interactions with humans' livestock and pets (e.g. cattle, pigs, goats), such as predation (in the case of vampire bats), an accidental encounter, or an animal scavenging a bat carcass, compound the risk of zoonotic transmission.[98]
Among ectoparasites, bats carry fleas and mites, as well as specific parasites called bat bugs.[99] However, they are one of the few mammalian orders that cannot host lice (most of the others are water animals). This may be due to overwhelming competition from more effective, specialized parasites, such as the bat bugs which occupy the same niche.[100]
They are also implicated in the emergence of SARS (severe acute respiratory syndrome), since they serve as a natural host for the type of virus involved (the genus Coronavirus, whose members typically cause mild respiratory disease in humans). A joint CAS/CSIRO team using phylogenetic analysis found that the SARS Coronavirus originated within the SARS-like Coronavirus group carried by the bat population in China.[101] However, note that they only served as the source of the precursor virus (which "jumped" to humans and evolved into the strain responsible for SARS): bats do not carry the SARS virus itself.[91]
Rabies
As of 2016, bats present a significant hazard in areas where the virus is endemic (such as the southern United States). They serve as the natural reservoirs for the rabies virus. For example, studies performed on Mexican free-tailed bats in Austin, Texas found an exposure rate of 45% among otherwise healthy individuals.[98]
In the United States, bats typically constitute around a quarter of reported cases of rabies in wild animals. However, their bites account for the vast majority of cases of rabies in humans.[102] Of the 36 cases of domestically acquired rabies recorded in the country in 1995–2010, two were caused by dog bites and four patients were infected by receiving transplants from an organ donor who had previously died of rabies. All other cases were caused by bat bites.[103]
Rabies is considered fully preventable if the patient is administered a vaccine prior to the onset of symptoms. However, unlike raccoon or skunk bites, bat bites may go ignored or unnoticed and hence untreated. Many victims may not realize they have been bitten, because bats have very small teeth and do not always leave obvious marks. Victims may also be bitten while sleeping or intoxicated, and children, pets, and the mentally handicapped are especially vulnerable.[104] Rabid bats are broadly distributed throughout the United States; in 2008–2010, cases were reported in every state except Alaska and Hawaii, and Puerto Rico.
The most severe threat to humans and domestic animals comes from sick, downed, or dead bats, which typically have a very high infection rate (e.g. 70% for the Austin bats).[98] Furthermore, since they may be clumsy, disoriented, and unable to fly, these stricken bats are much more likely to come into contact with humans.
Public health organizations such as the CDC generally recommend that any contact with a potentially infected animal (including any bat) be reported promptly, and those at risk of infection are treated with a post-exposure prophylaxis (PEP) regimen to prevent contraction of the virus, which is near-universally fatal with very few exceptions. 30,000 PEP treatments are performed each year in the US, in large part due to contact with bats.[91][98]
The Centers for Disease Control and Prevention (CDC) provide fully detailed information on all aspects of bat management in North America, including how to capture a bat, what to do in case of exposure, and how to bat-proof a house humanely.[105] In certain countries, such as the United Kingdom, it is illegal to handle bats without a license and advice should be sought from an expert organisation, such as the Bat Conservation Trust, if a trapped or injured bat is found. Where rabies is not endemic, as throughout most of Western Europe, small bats can be considered harmless. Larger bats may bite if handled.
There is evidence that bat rabies virus can infect victims purely through airborne transmission ("cryptic rabies"), without direct physical contact of the victim with the bat itself. This phenomenon has very rarely been reported, and has occurred among victims breathing virus-infected air in environments such as caves, after long exposures.[106][107]
Evidence suggests that all active widespread rabies strains (i.e. those affecting most terrestrial carnivores/omnivores) evolved from strains that were originally endemic to bats. Through zoonosis, these strains mutated and "jumped" to other species. In North America, for example, this jump reportedly occurred in the mid-1600s.[98]
Conservation efforts
Groups such as the Organization for Bat Conservation and Bat Conservation International aim to increase awareness of bats' ecological roles and the environmental threats they face.
In the United Kingdom, all bats are protected under the Wildlife and Countryside Acts, and even disturbing a bat or its roost can be punished with a heavy fine.
In Sarawak, Malaysia, bats are protected species under the Wildlife Protection Ordinance 1998 (see Malaysian Wildlife Law). The hairless bat (Cheiromeles torquatus) and Greater nectar bat are consumed by the local communities.
Bats can be a tourist attraction. The Congress Avenue Bridge in Austin, Texas is the summer home to North America's largest urban bat colony, an estimated 1,500,000 Mexican free-tailed bats, which eat an estimated 10,000 to 30,000 pounds of insects each night. An estimated 100,000 tourists per year visit the bridge at twilight to watch the bats leave the roost.[108]
Artificial roosts


Many people put up bat houses to attract bats just as some put up birdhouses to attract birds. Reasons for this vary, but most revolve around the bats being the primary nocturnal insectivores in most, if not all, ecologies. They can be built or bought, and plans and guidelines can be found online.[109]
Constructed in March 1991, the University of Florida bat house is the largest occupied artificial roost in the world. The bat house has approximately 300,000 residents, which can eat upwards of 2.5 billion insects (2,500 pounds or 1,100 kilograms) per night.[110] This started when a 1987 fire caused a colony of bats residing in Johnson Hall's attic to become homeless: they were forced to move to the James G. Pressly Stadium on the north side of campus, where the odor and the guano from this newly arrived colony did not please spectators. To remedy the situation, in September 1991 thousands of bats were captured and transported to a newly built bat house. However, they abandoned it within days, and it sat empty for 3 years. Finally, in 1995, the bats moved in permanently, and the colony has continued to grow. In 2009, part of the original bat house collapsed: a new "Bat Barn" was constructed next to it, and the interior of the original was rebuilt.[111]
In Britain, British hardened field defences of World War II have been converted to make roosts for bats. Pillboxes that are well dug-in and thick-walled are naturally damp and provide the stable thermal environment required by bats that would otherwise hibernate in caves. With a few minor modifications, suitable pillboxes can be converted to artificial caves for bats.[112][113]
Again in the UK, purpose-built bat houses are occasionally built when existing roosts are destroyed by developments, such as new roads; one such has been built associated with bat bridges on the new (2008) A38 Dobwalls bypass.[114] These have been subject to criticism for their high cost per animal: the Dobwalls installation cost £300,000 for an estimated 40 bats.[115]
Nectar-rich plants attracting moths
Besides mosquitoes, some species of bats also consume moths. Moths benefit from the availability of nectar-rich plants.[116] A study by Bat Conservation International claims that, by eating moths, bats save the global corn industry more than a million dollars a year.[117]
Threats
.jpg)
While conservation efforts are in place to protect bats, many threats still remain.
White nose syndrome
White nose syndrome is a condition associated with the deaths of millions of bats in the Eastern United States and Canada.[118] The disease is named after a white fungus, Pseudogymnoascus destructans, found growing on the muzzles, ears, and wings of afflicted bats. This fungus, which is mostly spread from bat to bat, is the sole cause of the disease.[119] The fungus was first discovered in central New York State in 2006 and spread quickly to the entire Eastern US north of Florida; mortality rates of 90–100% have been observed in most caves.[120] New England and the mid-Atlantic states have, since 2006, witnessed entire species completely extirpated and others with numbers that have gone from the hundreds of thousands, even millions, to a few hundred or less.[121] The provinces of Nova Scotia, Quebec, Ontario, and New Brunswick have witnessed identical die offs, with the Canadian government making preparations to protect all remaining bat populations in its territory.[122] Scientific evidence suggests that longer winters where the fungus has a longer period of time to infect bats results in greater chances of mortality.[123][124][125]
As of 2014, the geographical spread of white nose syndrome in North American bats has worsened in the sense that infection has crossed the Mississippi River.[126] Species native to northern Mexico and the West have not yet been affected, but it remains unknown how or if white nose syndrome will affect bat species in these regions.[127]
Barotrauma and wind turbines
Evidence suggests that barotrauma is causing bat fatalities around wind farms.[128] The lungs of bats are typical mammalian lungs, and unlike the lungs of birds, they are thought to be more sensitive to sudden air pressure changes in their immediate vicinity, such as near wind turbines, and are more liable to rupture.[129] Bats suffer a higher death rate than birds in the neighborhood of wind turbines.[130][131][132] Since there are no signs of external trauma, the cause has been hypothesized to be a greater sensitivity to sudden pressure fluctuations in the mammalian lung than in that of birds.[133] In addition, it has been suggested that bats are attracted to these structures, perhaps seeking roosts, and thereby increasing the death rate.[129] Acoustic deterrents may prove beneficial in mitigating bat mortality at wind energy facilities.[134]
Bats in human culture
Mythology

In European cultures, bats have long been associated with witchcraft, black magic and darkness. The Weird Sisters incorporate the fur of a bat in their brew in Shakespeare's Macbeth, written around 1603-1605.[135] Because bats are mammals, yet can fly, this gives them status as liminal beings in many cultural traditions.[136] In Western culture, the bat is often a symbol of the night and its foreboding nature. The bat is a primary animal associated with fictional characters of the night, both villains, such as Dracula, and heroes, such as Batman. The association of the fear of the night with the animal was treated as a literary challenge by Kenneth Oppel, who created a best-selling series of novels, beginning with Silverwing, which feature bats as the central heroic figures much as anthropomorphized rabbits were the central figures to the classic novel Watership Down.
Among some Native Americans, such as the Creek, Cherokee and Apache, the bat is a trickster spirit. In Tanzania, a winged bat cryptid known as Popobawa, is believed to be a shapeshifting evil spirit that assaults and sodomises its victims.[137]
Not all legends surrounding bats are negative. Chinese lore claims that the bat is a symbol of longevity and happiness, and the bat is similarly lucky in Poland, geographical Macedonia and among the Kwakiutl and Arabs.
An old wives' tale has it that bats will entangle themselves in people's hair. One likely source of this belief is that insect-eating bats seeking prey may dive erratically toward people, who attract mosquitoes and gnats, leading the squeamish to believe the bats are trying to get in their hair.
Aztec
In Mesoamerican mythology during the Classic-Contemporary period, bats symbolized the land of the dead, which was considered to be the underworld.[138] They also symbolized destruction and decay. Bats may have symbolized in this way because they fly only at night and dwell in caves during the daytime and are associated with human skulls and bones by classic Maya ceramists. Central Mexicans sometimes depicted bats having snouts that looked like "sacrificial knives and carrying human head" in the Postclassic era.[139] Bat images were engraved onto funerary urns, and were emphasized with large claws and round ears by Zapotecs. They were commonly associated with death.[140] The depiction of bats on funeral urns and goods took on some the characteristics of the jaguar, which was, and still is, another entity of the night and the underworld. There have also been instances where bats are portrayed next to other animals portrayed negatively in Mesoamerica, including scorpions and other nocturnal animals such as owls. Pre-Columbian cultures associated animals with gods, and often displayed them in art. The Moche people depicted bats in their ceramics.[141]
A life-size, ceramic bat-man was discovered and dug up from the Templo Mayor.[142] The Templo Mayor is located in the center of the Mexica capital of Tenochtitlan. Known as a god of death, this statue has the clawed feet and hands of a bat, but the body of a man. The statue's human-like eyes bulged out from the bat-like head, making the Zapotec images very realistic and living. In the 1930s, the Kaqchikel Maya were said to have proclaimed the bat was the Devil's provider. Kaqchikel would leave the Devil's underworld home and collect blood from the animals to be used for scrumptious meals to feed the Devil. "In the myths, the beast of prey and the animal that is preyed upon play two significant roles. They represent two aspects of life—the aggressive, killing, conquering, creating aspect of life, and the one that is the matter or, you might say, the subject matter".[143] In the Devil's underworld, dead sinners would work off their sins to get to heaven, indicating that the bat, too, was a sinner and worked under the authority of the Devil.[144]
Oaxaca

According to Oaxacan mythology, the bat's nocturnal nature can be traced back to its ancient jealousy of birds' feathers. One day, as the myth goes, the bat felt isolated and undesirable, and told God that he was cold. God, fair and just, turned to birds in the animal kingdom and asked if they would show compassion and donate a feather to the bat to help him keep warm. The birds all agreed and began to pluck one feather from their bodies to give to the bat. With all of these feathers, the bat became even more magnificent-looking than all birds, and was able to spread color to the night sky. During daylight, the bat created rainbows that reflected vibrant colors from the sun. With his new beauty and abilities, the bat soon became arrogant and conceited. The birds grew tired of the bat's self-glorification and decided to fly up to heaven and ask God to do something. When the birds told God of the bat's behaviour, He was surprised and decided to take a look Himself. When on earth, God called on the bat to show him what he was doing. The bat began to fly across the light blue sky where, one by one, each feather began to fall out, uncovering the bat's natural, ugly-looking body. When all his feathers were gone, the bat became distressed and ashamed of his appearance. He decided to hide in caves during the day and only come out during the night to search for his long-lost feathers.[144]
East Nigeria
According to a particular East Nigerian tale, the bat developed its nocturnal habits after causing the death of his partner, the bush-rat. The bat and the bush-rat would share activities, such as rummaging through the grass and trees, hunting, talking and bonding during the day. When at night, the bat and the bush-rat would alternate in cooking duties, cooking what was caught, and eat together. It appeared to be a dedicated partnership, but the bat hated the bush-rat immensely. The bush-rat always found the bat's soup more appetising, so when eating dinner one night, asked the bat why the soup tasted better than his own and also asked how it was made. The bat agreed to show him how to make it the next day, but instead was forming a malicious plan.
Next day, as the bat prepared his soup, the bush-rat came, greeting him and asking if he could be shown what was agreed yesterday. Earlier, the bat had found a pot looking exactly like the one he used usually, but it held warm water and so decided to use this instead. The bat explained to the bush-rat that to make his soup, he had to boil himself prior to serving the soup, where sweetness and flavor of the soup came from the flesh. The bat jumped in the pot seemingly excited, with the bush-rat mesmerised. After a few minutes, the bat climbed out and while the bush-rat was distracted, switched pots. The bat then served his soup out of the soup pot, both tasted it. Overanxious and eager, the bush-rat jumped into the pot of warm water. He stayed much longer in the pot, dying in the process.
When the bush-rat's wife returned that night to find her husband dead, she wept and ran to the chief of the land's house, telling him about what had happened and what she was sure the bat had done. In hearing this, the chief became angry, ordering for the immediate arrest of the bat. It just so happened that the bat was flying over the house and overheard what was just said. He quickly went into hiding high up in a tree. When the chief's men went looking for the bat, he could not be found. The search to arrest the bat carried on over several days, but he still could not be found. The bat needed to eat, so he flew out of hiding every night to hunt for food to avoid being arrested. This, according to Eastern Nigeria mythology, is why bats only fly at night.[145]
Tongan
The bat is sacred in Tonga and is often considered the physical manifestation of a separable soul.[146]
Heraldry

The bat is sometimes used as a heraldic symbol in Spain. The coats of arms of certain cities in eastern Spain, such as Valencia, Palma de Mallorca and Fraga, have the bat over the shield. Formerly, the Barcelona city coat of arms also had a bat crowning it, but the bat has been removed in the present-day versions. Heraldic use of the bat in Valencia, Catalonia and the Balearic Islands has its origins in a winged dragon (vibra or vibria), which featured in King James I of Aragon's helmet or cimera reial. This is the most widely accepted theory, although there is also a legend that says that, due to the intervention of a bat, King James was able to win a crucial battle against the Saracens that allowed him to win Valencia for his kingdom. The use of the bat as a heraldic symbol is prevalent in the territories of the former Crown of Aragon and it is little used elsewhere. However, it can be found in a few places, as in the coats of arms of the city of Albacete, in Spain, as well as the town of Montchauvet (Yvelines), in France.
Non heraldic organizations also use bats in their symbols. Certain Spanish football clubs including Valencia CF and Levante UD use bats in their badges. The burgee of the Royal Valencia Yacht Club (Reial Club Nàutic de València) displays a bat on a golden field in its center.[147]
Depictions in art
-
_-_Francisco_de_Goya_y_Lucientes.jpg)
Francisco Goya, The Sleep of Reason Produces Monsters, 1797
-

Biho Takashi, Bat Before the Moon, 1910, Brooklyn Museum
-

Crescent-Shaped Ornament with Bat. Moche, C.E. 1 – 300, Brooklyn Museum
State symbols
Three U.S. states have an official state bat. Texas and Oklahoma are represented by the Mexican free-tailed bat; Virginia is represented by the Virginia big-eared bat.[148]
See also
- Ann W. Richards Congress Avenue Bridge, location of the world's largest urban bat colony in Austin, Texas
- Arctic rabies virus
- Bat Conservation International
- Bat detector
- Bat (food) – bats are a food source for humans in some countries and cultures
- Grandview Mine, a bat-protection gating project in Grand Canyon National Park
- Organization for Bat Conservation
References
- Notes
- ↑ Ancient Greek: χείρ, Henry George Liddell, Robert Scott, A Greek-English Lexicon, on Perseus
- ↑ Ancient Greek: πτερόν, Henry George Liddell, Robert Scott, A Greek-English Lexicon, on Perseus
- 1 2
 Chisholm, Hugh, ed. (1911). "Chiroptera". Encyclopædia Britannica. 6 (11th ed.). Cambridge University Press. pp. 239–247.
Chisholm, Hugh, ed. (1911). "Chiroptera". Encyclopædia Britannica. 6 (11th ed.). Cambridge University Press. pp. 239–247. - 1 2 Hunter, P. (September 2007). "The nature of flight. The molecules and mechanics of flight in animals". Science and Society. 8 (9): 811–813. doi:10.1038/sj.embor.7401050. PMC 1973956
 . PMID 17767190. Retrieved 2009-07-17.
. PMID 17767190. Retrieved 2009-07-17. - ↑ Tudge, Colin (2000). The Variety of Life. Oxford University Press. ISBN 0-19-860426-2.
- ↑ "Bumblebee bat (Craseonycteris thonglongyai)". EDGE Species. Retrieved 2008-04-10.
- ↑ "Kitti's Hog-Nosed Bat: Craseonycteridae - Physical Characteristics - Bats, Bumblebee, Species, Inches, Brown, and Tips". Animals.jrank.org. Retrieved 2013-06-24.
- ↑ Wood, Gerald (1983). The Guinness Book of Animal Facts and Feats. ISBN 978-0-85112-235-9.
- ↑ Nowak, R. M., editor (1999). Walker's Mammals of the World. Vol. 1. 6th edition. Pp. 264–271. ISBN 0-8018-5789-9
- ↑ http://phys.org/news/2016-11-brazilian-free-tailed-fastest-flyer-animal.html
- ↑ "Bat, noun 2". Online Etymology Dictionary. Retrieved 2013-06-24.
- ↑ Dave Smith. "Chiroptera: Systematics".
- ↑ Pumo, D.E.; et al. (1998). "Complete Mitochondrial Genome of a Neotropical Fruit Bat, Artibeus jamaicensis, and a New Hypothesis of the Relationships of Bats to Other Eutherian Mammals". Journal of Molecular Evolution. 47 (6): 709–717. doi:10.1007/PL00006430. PMID 9847413.
- ↑ Zhou, X.; et al. (2011). "Phylogenomic Analysis Resolves the Interordinal Relationships and Rapid Diversification of the Laurasiatherian Mammals". Systematic Biology. 61 (1): 150–164. doi:10.1093/sysbio/syr089. PMC 3243735. PMID 21900649.
- ↑ Holland, Richard A.; et al. (Dec 2004). "Echolocation signal structure in the Megachiropteran bat Rousettus aegyptiacus Geoffroy 1810". J Exp Biol. 207 (25): 4361–4369. doi:10.1242/jeb.01288.
- ↑ Brown, William M. (September 2001). "Natural selection of mammalian brain components". Trends in Ecology and Evolution. 16 (9): 471–473. doi:10.1016/S0169-5347(01)02246-7.
- 1 2 James Stephen; Olney, Peter (1994). Creative Conservation: Interactive Management of Wild and Captive Animals. Springer. p. 352. ISBN 9780412495700.
- 1 2 3 4 Nancy B. Simmons1, Kevin L. Seymour2, Jo¨rg Habersetzer3 & Gregg F. Gunnell4 (2008). "Primitive Early Eocene bat from Wyoming and the evolution of flight and echolocation". Nature. 451 (7180): 818–21. Bibcode:2008Natur.451..818S. doi:10.1038/nature06549. PMID 18270539.
- ↑ "Primitive Early Eocene bat from Wyoming and the evolution of flight and echolocation". Nature. 2008-02-14. Retrieved 2008-07-03.
- ↑ Pettigrew JD, Maseko BC, Manger PR (April 2008). "Primate-like retinotectal decussation in an echolocating megabat, Rousettus aegyptiacus". Neuroscience. 153 (1): 226–31. doi:10.1016/j.neuroscience.2008.02.019. PMID 18367343.
- 1 2 3 Tsagkogeorga, G; Parker, J; Stupka, E; Cotton, JA; Rossiter, SJ (2013). "Phylogenomic analyses elucidate the evolutionary relationships of bats (Chiroptera)". Current Biology. 23 (22): 2262–2267. doi:10.1016/j.cub.2013.09.014. PMID 24184098.
- 1 2 Teeling, E.C.; Springer, M. S.; Madsen, O.; Bates, P.; O'Brien, S. J.; Murphy, W. J. (2005). "A Molecular Phylogeny for Bats Illuminates Biogeography and the Fossil Record". Science. 307 (5709): 580–584. Bibcode:2005Sci...307..580T. doi:10.1126/science.1105113. PMID 15681385.
- ↑ Teeling; Teeling, Emma C.; Scally, Mark; Kao, Diana J.; Romagnoli, Michael L.; Springer, Mark S.; et al. (2000). "Molecular evidence regarding the origin of echolocation and flight in bats". Nature. 403 (6766): 188–192. Bibcode:2000Natur.403..188T. doi:10.1038/35003188. PMID 10646602.
- ↑ "Order Chiroptera (Bats)". Animal Diversity Web. Archived from the original on 21 December 2007. Retrieved 2007-12-30.
- ↑ Springer; Teeling, E. C.; Madsen, O.; Stanhope, M. J.; De Jong, W. W.; et al. (2001). "Integrated fossil and molecular data reconstruct bat echolocation". Proceedings of the National Academy of Sciences. 98 (11): 6241–6246. Bibcode:2001PNAS...98.6241S. doi:10.1073/pnas.111551998. PMC 33452. PMID 11353869.
- ↑ Li et al. 2007. "Accelerated FoxP2 Evolution in Echolocating Bats". PLOS ONE.
- ↑ Li et al. 2008. "The hearing gene Prestin reunites the echolocating bats". Proc. Natl. Academy. Sci. U.S.A.
- ↑ James M. Hutcheon & John A.W. Kirsch. "A moveable face: deconstructing the Microchiroptera and a new classification of extant bats" (PDF). doi:10.3161/1733-5329(2006)8%5B1:AMFDTM%5D2.0.CO;2. Archived from the original (PDF) on 2006-08-22.
- ↑ Eick; Jacobs, DS; Matthee, CA; et al. (2005). "A Nuclear DNA Phylogenetic Perspective on the Evolution of Echolocation and Historical Biogeography of Extant Bats (Chiroptera)". Molecular Biology and Evolution. 22 (9): 1869–86. doi:10.1093/molbev/msi180. PMID 15930153.
Following the recommendations of Hutcheon and Kirsch (2004), we refer to the two suborders of chiropterans as 'Pteropodiformes' (comprising the Pteropodidae, Rhinolophidae, Hipposideridae, Megadermatidae, and Rhinopomatidae) and 'Vespertilioniformes' (remaining microbat families).
- ↑ Eick; Jacobs, DS; Matthee, CA; et al. (2005). "A Nuclear DNA Phylogenetic Perspective on the Evolution of Echolocation and Historical Biogeography of Extant Bats (Chiroptera)". Molecular Biology and Evolution. 22 (9): 1869–86. doi:10.1093/molbev/msi180. PMID 15930153.
Several molecular studies have shown that Chiroptera belong to the Laurasiatheria (represented by carnivores, pangolins, cetartiodactyls, eulipotyphlans, and perissodactyls) and are only distantly related to dermopterans, scandentians, and primates (Nikaido et al. 2000; Lin and Penny 2001; Madsen et al. 2001; Murphy et al. 2001a, 2001b; Van Den Bussche and Hoofer 2004).
- ↑ Zhang, G.; Cowled, C.; Shi, Z.; Huang, Z.; Bishop-Lilly, K. A.; Fang, X.; Wynne, J. W.; Xiong, Z.; Baker, M. L.; Zhao, W.; Tachedjian, M.; Zhu, Y.; Zhou, P.; Jiang, X.; Ng, J.; Yang, L.; Wu, L.; Xiao, J.; Feng, Y.; Chen, Y.; Sun, X.; Zhang, Y.; Marsh, G. A.; Crameri, G.; Broder, C. C.; Frey, K. G.; Wang, L.-F.; Wang, J. (2012). "Comparative Analysis of Bat Genomes Provides Insight into the Evolution of Flight and Immunity". Science. 339 (6118): 456–460. Bibcode:2013Sci...339..456Z. doi:10.1126/science.1230835.
- ↑ Eiting, T.P.; Gunnell, G.F. (2009). "Global completeness of the bat fossil record". Journal of Mammalian Evolution. 16: 151–173. doi:10.1007/s10914-009-9118-x.
- ↑ Simmons, N.B., K.L. Seymour, J. Habersetzer, and G.F. Gunnell. 2008. Primitive early Eocene bat from Wyoming and the evolution of flight and echolocation" Nature 451:818-U816.
- ↑ (BBC News) "Bat fossil solves evolution poser" 13 February 2008.
- ↑ (Discovery Channel article) "Prehistoric bats learned to fly before they could see" February 13, 2008.
- ↑ Bishop, K.L. (2008). "The Evolution of Flight in Bats: Narrowing the Field of Plausible Hypotheses". The Quarterly Review of Biology. 83 (2): 153–169. doi:10.1086/587825. PMID 18605533.
- ↑ Paleontologists Determine Original Color of Extinct Bats
- ↑ Thomas, S.P., and R.A. Suthers. 1972. Physiology and energetics of bat flight" Journal of Experimental Biology 57:317-&.
- ↑ "Bats of the World". Bat Conservation Trust. Archived from the original on 5 January 2011. Retrieved January 2011. Check date values in:
|access-date=(help) - ↑ Grzimek's Animal Life Encyclopedia: Vol 13 Mammals II (2nd ed.). 2003. p. 311. ISBN 0-7876-5362-4.
- ↑ "Bat Conservation International: Species Profiles".
- ↑ "The Mysterious White Nose Syndrome and the 10 Bat Species Most at Risk".
- ↑ "California Bats".
- 1 2 Sears, K. E.; Behringer, R. R.; Rasweiler, J. J.; Niswander, L. A. (2006). "Development of bat flight: Morphologic and molecular evolution of bat wing digits". Proceedings of the National Academy of Sciences. 103 (17): 6581–6586. Bibcode:2006PNAS..103.6581S. doi:10.1073/pnas.0509716103. PMC 1458926. PMID 16618938.
- ↑ Bats In Flight Reveal Unexpected Aerodynamics
- ↑ Bats save energy by drawing in wings on upstroke
- 1 2 Roberts, W.C. (October 2006). "Facts and ideas from anywhere". Proceedings (Baylor University. Medical Center). 19 (4): 425–434. PMC 1618737. PMID 17106509. Retrieved 2009-07-17.
- ↑ Irwin, N. (March 1997). "Wanted DNA samples from Nyctimene or Paranyctimene Bats" (PDF). The New Guinea Tropical Ecology and Biodiversity Digest. 3: 10. Retrieved 2009-07-17.
- 1 2 3 Melissa Calhoun (15 December 2005). "Bats Use Touch Receptors on Wings to Fly, Catch Prey, Study Finds". Archived from the original on 7 September 2006. Retrieved 2006-10-18.
- ↑ Jones, T. Wharton (1852-01-01). "Discovery That the Veins of the Bat's Wing (Which are Furnished with Valves) are Endowed with Rythmical Contractility, and That the Onward Flow of Blood is Accelerated by Each Contraction". Philosophical Transactions of the Royal Society of London. 142: 131–136. JSTOR 108539.
- ↑ Dongaonkar, Ranjeet M.; Quick, Christopher M.; Vo, Jonathan C.; Meisner, Joshua K.; Laine, Glen A.; Davis, Michael J.; Stewart, Randolph H. (2012-06-15). "Blood flow augmentation by intrinsic venular contraction in vivo". American Journal of Physiology - Regulatory, Integrative and Comparative Physiology. 302 (12): R1436–R1442. doi:10.1152/ajpregu.00635.2011. ISSN 0363-6119. PMC 3378342. PMID 22513742.
- ↑ Neuweiler, Gerhard (2000-01-01). The Biology of Bats. Oxford University Press. ISBN 9780195099515.
- ↑ Chamberlain, Ted (2006-12-06). "Photo in the News: Bat Has Longest Tongue of Any Mammal". National Geographic News. National Geographic Society. Archived from the original on 6 June 2007. Retrieved 2007-06-18.
A. fistulata (shown lapping sugar water from a tube) has the longest tongue, relative to body length, of any mammal—and now scientists think they know why.
- ↑ Maina, J. N. (2000-10-15). "What it takes to fly: the structural and functional respiratory refinements in birds and bats". Journal of Experimental Biology. 203 (20): 3045–3064. ISSN 0022-0949. PMID 11003817.
- ↑ "Bats and Roadside Mammals Survey Sonogram Analysis" (PDF). p. 10. Archived from the original (PDF) on 2007-10-21.
- ↑ Jones, K. E.; O. R. P. Bininda-Emonds & J. L. Gittleman (2005). "Bats, clocks, and rocks: diversification patterns in chiroptera". Evolution. 59 (10): 2243–2255. doi:10.1554/04-635.1. PMID 16405167.
- ↑ Teeling, E. C. (2009). "Hear, hear: the convergent evolution of echolocation in bats?". Trends in Ecology & Evolution. 24 (7): 351–354. doi:10.1016/J.Tree.2009.02.012. PMID 19482373.
- ↑ Jones, G. & M. W. Holderied (2007). "Bat echolocation calls: adaptation and convergent evolution". Proceedings of the Royal Society B: Biological Sciences. 274 (1612): 905–912. doi:10.1098/Rspb.2006.0200. PMC 1919403. PMID 17251105.
- ↑ DesRoche, K. M. B. Fenton & W. C. Lancaster (2007). "Echolocation and the thoracic skeletons of bats: a comparative morphological study". Acta Chiropterologica. 9 (2): 483–494. doi:10.3161/1733-5329(2007)9[483:EATTSO]2.0.CO;2. ISSN 1733-5329.
- ↑ Pavey, C. R.; Burwell, C. J. (1998). "Bat Predation on Eared Moths: A Test of the Allotonic Frequency Hypothesis". Oikos. 81 (1): 143–151. doi:10.2307/3546476. JSTOR 3546476.
- ↑ "The Bat's Ear as a Diffraction Grating". Oai.dtic.mil. Retrieved 2013-06-24.
- ↑ Kuc, Roman (May 2009). "Model predicts bat pinna ridges focus high frequencies to form narrow sensitivity beams". J. Acoust. Soc. Am. 125 (5): 3454–9. Bibcode:2009ASAJ..125.3454K. doi:10.1121/1.3097500. PMID 19425684.
- ↑ Surlykke, A.; Ghose, K.; Moss, C. F. (April 2009). "Acoustic scanning of natural scenes by echolocation in the big brown bat, Eptesicus fuscus". J. Exp. Biol. 212 (Pt 7): 1011–20. doi:10.1242/jeb.024620. PMC 2726860. PMID 19282498.
- ↑ Nida Sophasarun. "Experts debunk bats' bad rap". Online extra. National Geographic. Retrieved April 30, 2013.
- ↑ Rachel Kaufman (December 15, 2010). "Bats Crash More When They Rely on Vision". National Geographic Daily News. Retrieved April 30, 2013.
- ↑ "Smithsonian Institution". Si.edu. 2010-12-07. Retrieved 2013-06-24.
- ↑ Fenton, M. Brock (2001). Bats. New York: Checkmark Books. pp. 60–62. ISBN 0-8160-4358-2.
- 1 2 Fenton, M. Brock (2001). Bats. New York: Checkmark Books. pp. 93=94. ISBN 0-8160-4358-2.
- 1 2 Fenton, M. Brock (2001). Bats. New York: Checkmark Books. pp. 95–107. ISBN 0-8160-4358-2.
- 1 2 Fenton, M. Brock (2001). Bats. New York: Checkmark Books. pp. 4–5. ISBN 0-8160-4358-2.
- ↑ Nyffeler, Martin; Knörnschild, Mirjam (March 13, 2013). Bilde, Trine, ed. "Bat Predation by Spiders". Plos One. 3. 8 (3): e58120. Bibcode:2013PLoSO...858120N. doi:10.1371/journal.pone.0058120.
- ↑ Neuweiler, Gerhard (2000). Biology of Bats. ISBN 978-0-19-509950-8.
- ↑ "Rescue, Rehab, Release & Sanctuary". Archived from the original on 2006-10-10. Retrieved 22 October 2006.
- ↑ Gerald S. Wilkinson & Jason M. South (2002). "Life history, ecology and longevity in bats" (PDF). Aging Cell. Retrieved 22 Dec 2015.
- 1 2 3 4 5 6 7 8 Wilson, Don. Bats in Question. London: Smithsonian Institution Press, 1997
- 1 2 3 4 5 Lauber, Patricia. Bats: Wings in the Night. New York: Random House, 1968
- ↑ Stuart, Dee (1994). Bats: Mysterious Flyers of the Night (illustrated ed.). Lerner Publications. p. 18. ISBN 978-0-87614-814-3. Extract of page 18
- 1 2 Johnson, Sylvia. Bats. Minneapolis: Lerner Publications Company, 1985
- 1 2 3 4 5 6 Shebar, Sharon. Bats. New York: Franklin Watts, 1990
- ↑ Cui J, Yuan X, Wang L, Jones G, Zhang S (Nov 2011). "Recent loss of vitamin C biosynthesis ability in bats". PLoS ONE. 6 (11): e27114. Bibcode:2011PLoSO...627114C. doi:10.1371/journal.pone.0027114. PMC 3206078. PMID 22069493.
- ↑ A trace of GLO was detected in only one of 34 bat species tested, across the range of six families of bats tested: See Jenness, R., E. Birney, and K. Ayaz. 1980. Variation of L-gulonolactone oxidase activity in placental mammals. Comparative Biochemistry and Physiology 67B:195-204. Earlier reports of only fruit bats being deficient were based on smaller samples.
- ↑ Cui J, Pan YH, Zhang Y, Jones G, Zhang S (Feb 2011). "Progressive pseudogenization: vitamin C synthesis and its loss in bats". Mol. Biol. Evol. 28 (2): 1025–31. doi:10.1093/molbev/msq286. PMID 21037206.
- ↑ Fitt, G.P. (1989). "The ecology of Heliothis species in relation to agro-ecosystems". Annual Review of Entomology. 34: 17–52. doi:10.1146/annurev.ento.34.1.17.
- ↑ Arita, Hector T.; Santos-Del-Prado, Karina; Arita, Hector T. (1999). "Conservation Biology of Nectar-Feeding Bats in Mexico". Journal of Mammalogy. 80 (1): 31–41. doi:10.2307/1383205.
- ↑ Gerardo, H.; Hobson, Keith A.; Adriana, M. A; Daniel, E. B; Sanchez-Corero, Victor; German, M. C. (2001). "The Role of Fruits and Insects in the Nutrition of Frugivorous Bats: Evaluating the Use of Stable Isotope Models". Biotropica. 33 (3): 520–28. doi:10.1111/j.1744-7429.2001.tb00206.x.
- ↑ Hodgkison, Robert; Balding, Sharon T.; Zuibad, Akbar; Kunz, Thomas H. (2003). "Fruit Bats (Chiroptera: Pteropodidae) as Seed Dispersers and Pollinators in a Lowland Malaysian Rain Forest". Biotropica. 35 (4): 491–502. doi:10.1111/j.1744-7429.2003.tb00606.x.
- ↑ Greenhall, Arthur M. 1961. Bats in Agriculture. A Ministry of Agriculture Publication. Trinidad and Tobago
- ↑ Jones, V. (2000). "Drinking in the river". Vivian Jones. Archived from the original on 11 June 2009. Retrieved 17 July 2009.
- ↑ Wong, Samson; Susanna Lau; Patrick Woo; Kwok-Yung Yuen (2006-10-16). "Bats as a continuing source of emerging infections in humans" (Review). Reviews in Medical Virology. John Wiley & Sons. 17 (2): 67–91. doi:10.1002/rmv.520. PMID 17042030. Retrieved 2007-12-29.
The currently known viruses that have been found in bats are reviewed and the risks of transmission to humans are highlighted. (from abstract)
- ↑ McColl, KA; N Tordo; AA Aquilar Setien (April 2000). "Bat lyssavirus infections". Revue scientifique et technique. 19 (1): 177–196. PMID 11189715.
Bats, which represent approximately 24% of all known mammalian species, frequently act as vectors of lyssaviruses. (from abstract)
- 1 2 3 "CDC Features - Take Caution When Bats Are Near". www.cdc.gov. Retrieved 2016-08-21.
- ↑ Halpin, K.; P. L. Young; H. E. Field; J. S. Mackenzie (August 1, 2000). "Isolation of Hendra virus from pteropid bats: a natural reservoir of Hendra virus". Journal of General Virology. 81 (8): 1927–1932. PMID 10900029. Retrieved 2007-12-29.
In this paper we describe the isolation of HeV from pteropid bats, corroborating our serological and epidemiological evidence that these animals are a natural reservoir host of this virus.
- ↑ Leroy, Eric M.; Brice Kumulungui; Xavier Pourrut; Pierre Rouque; et al. (2005-12-01). "Fruit bats as reservoirs of Ebola virus" (Brief Communication). Nature. 438 (7068): 575–576. Bibcode:2005Natur.438..575L. doi:10.1038/438575a. PMID 16319873. Retrieved 2007-12-29.
We find evidence of asymptomatic infection by Ebola virus in three species of fruit bat, indicating that these animals may be acting as a reservoir for this deadly virus. (from abstract)
- ↑ Charles Q. Choi (March 2006). "Going to Bat". Scientific American. pp. 24, 26. Retrieved 2007-12-29.
Long known as vectors for rabies, bats may be the origin of some of the most deadly emerging viruses, including SARS, Ebola, Nipah, Hendra and Marburg.
Note: This could be considered a lay summary of the various scientific publications cited in the preceding sentence. - 1 2 "Bats Host More Than 60 Human-Infecting Viruses". Retrieved 2016-08-21.
- ↑ Dobson, Andrew P. (2005-10-28). "What Links Bats to Emerging Infectious Diseases?". Science. 310 (5748): 628–629. doi:10.1126/science.1120872. ISSN 0036-8075. PMID 16254175.
- ↑ "Why Do Bats Transmit So Many Diseases?". Retrieved 2016-08-21.
- 1 2 3 4 5 Calisher, Charles H.; Childs, James E.; Field, Hume E.; Holmes, Kathryn V.; Schountz, Tony (2006-07-01). "Bats: Important Reservoir Hosts of Emerging Viruses". Clinical Microbiology Reviews. 19 (3): 531–545. doi:10.1128/CMR.00017-06. ISSN 0893-8512. PMC 1539106. PMID 16847084.
- ↑ Klimpel, Sven; Mehlhorn, Heinz (2013-10-10). Bats (Chiroptera) as Vectors of Diseases and Parasites: Facts and Myths. Springer Science & Business Media. ISBN 9783642393334.
- ↑ Clayton, Dale H.; Bush, Sarah E.; Johnson, Kevin P. (2015-12-24). Coevolution of Life on Hosts: Integrating Ecology and History. University of Chicago Press. ISBN 9780226302270.
- ↑ Li, Wendong; Z. Shi; M. Yu; W. Ren; et al. (2005-10-28). "Bats are natural reservoirs of SARS-like coronaviruses". Science. 310 (5748): 676–679. Bibcode:2005Sci...310..676L. doi:10.1126/science.1118391. PMID 16195424. Retrieved 2007-12-29. Lay summary – Science (2005-10-28).
The genetic diversity of bat-derived sequences supports the notion that bats are a natural reservoir host of the SARS cluster of coronaviruses.
- ↑ Bruessow, H. (2012=. On Viruses, Bats and Men: A Natural History of Food-Borne Viral Infections. In: Witzany G (ed). Viruses: Essential Agents of life. Springer, 245-267. doi:10.1007/978-94-007-4899-6_12
- ↑ "Center for Disease Control and Prevention: Human Rabies".
- ↑ "CDC: Learning about Bats and Rabies". CDC.gov. 22 April 2011. Retrieved 21 August 2016.
- ↑ "CDC's website on bats and rabies". Cdc.gov. 2010-03-22. Retrieved 2013-06-24.
- ↑ Constantine, Denny G. (April 1962). "Rabies transmission by nonbite route". Public Health Reports. Public Health Service. 77 (4): 287–289. doi:10.2307/4591470. PMC 1914752. PMID 13880956.
These findings support consideration of an airborne medium, such as an aerosol, as the mechanism of rabies transmission in this instance.
- ↑ Messenger, Sharon L.; Jean S. Smith; Charles E. Rupprecht (2002-09-15). "Emerging Epidemiology of Bat-Associated Cryptic Cases of Rabies in Humans in the United States". Clinical Infectious Diseases. 35 (6): 738–747. doi:10.1086/342387. PMID 12203172. Retrieved 2007-12-29.
Cryptic rabies cases are those in which a clear history of exposure to rabies virus cannot be documented, despite extensive case-history investigation. Absence of a documented bite history reflects inherent difficulties in obtaining accurate animal-contact information ... Thus, absence of bite-history data does not mean that a bite did not occur.
- ↑ FOX. "Best time to see the bat colony emerge from Congress Bridge in Downtown Austin". Retrieved 2016-08-21.
- ↑ "Bat Conservation International". Web.archive.org. 2002-01-24. Archived from the original on June 23, 2013. Retrieved 2013-06-24.
- ↑ "UF Bat Colony :: Florida Museum of Natural History". www.flmnh.ufl.edu. Retrieved 2016-08-21.
- ↑ Glover, Martin. "Facts about this colony". UF News. University of Florida. p. 1. Retrieved 2012-03-16.
.
- ↑ "Protecting and managing underground sites for bats (pdf), see section 6.4." (PDF). Retrieved 2006-05-18.
- ↑ "Pillbox converted to bat retreat, BBC website". BBC News. 2006-04-06. Retrieved 2006-05-18.
- ↑ "Bypass wings it with bat bridges". BBC. 2008-04-04. Retrieved 2016-08-21.
- ↑ "Bat bridges cost £27k per animal". BBC. 2009-10-22. Retrieved 2016-08-21.
- ↑ Pauline Pears (2005), HDRA encyclopedia of organic gardening, Dorling Kindersley, ISBN 978-1405308915
- ↑ 'Bats with Benefits', article in National Geographic, July 2016.
- ↑ "White-Nose Syndrome (WNS)". National Wildlife Health Center, U.S. Geological Survey. Retrieved 3 June 2014.
- ↑ "Culprit Identified: Fungus Causes Deadly Bat Disease". United States Geological Survey. 2011-10-26. Retrieved 2012-05-05.
- ↑ "White-Nose Syndrome - Background". Canadian Cooperative Wildlife Health Centre. Retrieved 3 June 2014.
- ↑ Daly M (14 November 2013). "Pennsylvania's Bats Nearly Wiped Out". CBS Philadelphia. Retrieved 21 April 2016.
- ↑ Gutenberg G (7 June 2012). "White-nose syndrome killing Canada's bats". Postmedia Network Inc. Retrieved 21 April 2016.
- ↑ "Canada : Environment Canada Announces Funding to Fight Threat of White-nose Syndrome to Bats.". Mena Report. Apr 6, 2013. Retrieved 3 June 2014.
- ↑ "Social Bats Pay a Price: Fungal Disease, White-Nose Syndrome ... Extinction?". The National Science Foundation. July 3, 2012. Retrieved 3 June 2014.
- ↑ Frick, W. F.; Pollock, J. F.; Hicks, A. C.; Langwig, K. E.; Reynolds, D. S.; Turner, G. G.; Butchkoski, C. M.; Kunz, T. H. (5 August 2010). "An Emerging Disease Causes Regional Population Collapse of a Common North American Bat Species". Science 329 (5992): 679–682. doi:10.1126/science.1188594. PMID 20689016.
- ↑ "White-Nose Syndrome Confirmed in Illinois Bats: Illinois becomes 20th state in U.S. to confirm deadly disease in bats" (PDF). Illinois Department of Natural Resources. 28 February 2013.
- ↑ "As White Nose Syndrome Spreads, Worries Persist About Potential Impact on Bats, Ag Industry". The Public Radio Service of Western Kentucky University. May 29, 2014. Retrieved 3 June 2014.
- ↑ Erin F. Baerwald et al, Barotrauma is a significant cause of bat fatalities at wind turbines Current Biology, 2008
- 1 2 "B.C. study to help bats survive wind farms", National Wind Watch, September 23, 2008, retrieved 19 April 2015
- ↑ "Bats take a battering at wind farms", New Scientist, May 12, 2007
- ↑ "Caution Regarding Placement of Wind Turbines on Wooded Ridge Tops" (PDF). Bat Conservation International. 4 January 2005. Archived (PDF) from the original on 23 May 2006. Retrieved 2006-04-21.
- ↑ Arnett, Edward B.; Wallace P. Erickson; Jessica Kerns; Jason Horn (June 2005). "Relationships between Bats and Wind Turbines in Pennsylvania and West Virginia: An Assessment of Fatality Search Protocols, Patterns of Fatality, and Behavioral Interactions with Wind Turbines" (PDF). Bat Conservation International. Archived from the original (PDF) on 2006-02-10. Retrieved 2006-04-21.
- ↑ Baerwald, Erin F; D'Amours, Genevieve H; Klug, Brandon J; Barclay, Robert MR (2008-08-26). "Barotrauma is a significant cause of bat fatalities at wind turbines". Current Biology. 18 (16): R695–R696. doi:10.1016/j.cub.2008.06.029. OCLC 252616082. PMID 18727900. Lay summary – CBC Radio – Quirks & Quarks (2008-09-20). Laysource includes audio podcast of interview with author.
- ↑ Johnson, Joshua B. et al. (2012). Effects of Acoustic Deterrents on Foraging Bats. Newtown Square, PA: United States Department of Agriculture, U.S. Forest Service, Northern Research Station.
- ↑ de Vries, Ad (1976). Dictionary of Symbols and Imagery. Amsterdam: North-Holland Publishing Company. p. 36. ISBN 0-7204-8021-3.
- ↑ McCracken, Gary F. (1993). "Folklore and the Origin of Bats". BATS Magazine. Bats in Folklore. 11 (4).
- ↑ Saleh, Ally (19 July 2001). "Sex-mad 'ghost' scares Zanzibaris". BBC News. Zanzibar. Retrieved 29 December 2014.
- ↑ "Aztec Symbols". Aztec-history.net. Retrieved 2013-06-24.
- ↑ Kay Almere Read and Jason J. Gonzalez. 2000. Mesoamerican Mythology. Oxford University Press. pp. 132
- ↑ "Artists Inspired by Oaxaca Folklore Myths and Legends". Oaxacanwoodcarving.com. Retrieved 2013-06-24.
- ↑ Berrin, Katherine & Larco Museum. The Spirit of Ancient Peru:Treasures from the Museo Arqueológico Rafael Larco Herrera. New York: Thames and Hudson, 1997.
- ↑ Read, Kay Almere & Gonzalez, Jason J. (2002). Mesoamerican Mythology: A Guide to the Gods, Heroes, Rituals, and Beliefs of Mexico and Central America. Oxford University Press. p. 132. ISBN 9780195149098.
- ↑ Joseph Cambell and Bill Moyers. 1988. The Power of Myth. Doubleday. pp. 91
- 1 2 Kay Almere Read and Jason J. Gonzalez. 2000. Mesoamerican Mythology. Oxford University Press. pp. 132–134
- ↑ Arnott, Kathleen. 1962. African Myths and Legends. Oxford University Press. Pp. 150–152
- ↑ Grant, Gilbert S. "Kingdom of Tonga: Safe Haven for Flying Foxes". Batcon.org. Retrieved 2013-06-24.
- ↑ "El Club de Mar acoge el I Trofeo Cluma de la clase Optimist - Revista Barcos a Vela" (in Spanish). 2014-06-20. Retrieved 2016-08-21.
- ↑ "Official state bats". netstate.com. NSTATE, LLC. Archived from the original on March 8, 2008. Retrieved February 13, 2011.
- Bibliography
- Altringham, J.D. 1998. Bats: Biology and Behaviour. Oxford: Oxford University Press.
- Dobat, K.; Holle, T.P. 1985. Blüten und Fledermäuse: Bestäubung durch Fledermäuse und Flughunde (Chiropterophilie). Frankfurt am Main: W. Kramer & Co. Druckerei.
- Fenton, M.B. 1985. Communication in the Chiroptera. Bloomington: Indiana University Press.
- Findley, J.S. 1995. Bats: a Community Perspective. Cambridge: Press Syndicate of the University of Cambridge.
- Fleming, T.H. 1988. The Short-Tailed Fruit Bat: a Study in Plant-Animal Interactions. Chicago: The University of Chicago Press.
- Greenhall, Arthur H. 1961. Bats in Agriculture. A Ministry of Agriculture Publication. Trinidad and Tobago.
- Klimpel, S.; Mehlhorn, H. 2014. Bats (Chiroptera) as Vectors of Diseases and Parasites: Facts and Myths. Heidelberg New York Dordrecht London: Springer.
- Kunz, T.H. 1982. Ecology of Bats. New York: Plenum Press.
- Kunz, T.H.; Racey, P.A. 1999. Bat Biology and Conservation. Washington: Smithsonian Institution Press.
- Kunz, T.H.; Fenton, M.B. 2003. Bat Ecology. Chicago: The University of Chicago Press.
- Neuweiler, G. 1993. Biologie der Fledermäuse. Stuttgart: Georg Thieme Verlag.
- Nowak, Ronald M. 1994. Walker's Bats of the World. Baltimore: The Johns Hopkins University Press.
- John D. Pettigrew's summary on Flying Primate Hypothesis
- Richarz, K. & Limbruner, A. 1993. The World of Bats. Neptune City: TFH Publications.
- Teeling, E.C. 2009. Chiroptera. Oxford University Press.
- Twilton, B. 1999. My Life as The Bat. Liverpool Hope University Press.
Further reading
- Davis, William B.; David J. Schmidly (1997) [1947]. "Order Chiroptera:Bats". In Lisa Bradley. The Mammals of Texas – Online Edition. Dave Scarbrough (1994 ed.). Texas Parks and Wildlife Department. Retrieved 2008-07-10.
External links
| The Wikibook Dichotomous Key has a page on the topic of: Chiroptera |
 Data related to Chiroptera at Wikispecies
Data related to Chiroptera at Wikispecies- Basic facts about bats
- UK Bat Conservation Trust
- University of Michigan Museum of Zoology
- Tree of Life
- Microbat Vision
- Bat Conservation International (US)
- Organization for Bat Conservation (US)
- Evolution of cranial morphology in bats in relation to diet (by Divulgare)
- The DSP Behind Bat Echolocation - Analysis of several kinds of bat echolocation