Posterior cingulate
| Posterior cingulate cortex | |
|---|---|
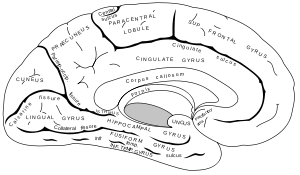 Medial surface of left cerebral hemisphere. | |
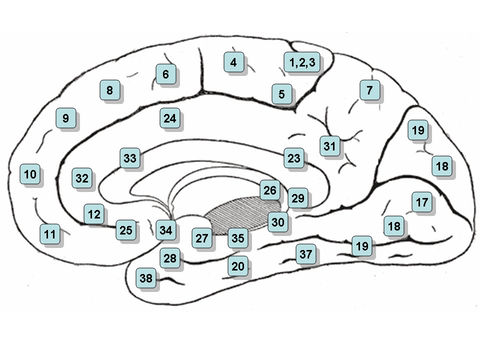 Medial surface. (Areas 23 and 31 at center right. Image is reversed from image above.) | |
| Details | |
| Part of | Cingulate gyrus |
| Identifiers | |
| Latin | Cortex cingularis posterior |
| NeuroNames | hier-144 |
| NeuroLex ID | Posterior cingulate cortex |
| FMA | 61924 |
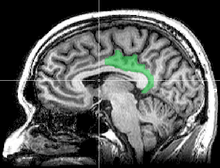
The posterior cingulate cortex (PCC) is the backmost part of the cingulate cortex, lying behind the anterior cingulate cortex. This is the upper part of the "limbic lobe". The cingulate cortex is made up of an area around the midline of the brain. Surrounding areas include the retrosplenial cortex and the precuneus.
Cytoarchitectonically the posterior cingulate cortex is associated with Brodmann areas 23 and 31.
The posterior cingulate cortex forms a central node in the default mode network of the brain. It has been shown to communicate with various brain networks simultaneously and is involved in various functions.[1] Along with the precuneus, the posterior cingulate cortex has been implicated as a neural substrate for human awareness in numerous studies of both the anesthesized and vegetative (coma) state. Imaging studies indicate a prominent role for the posterior cingulate cortex in pain and episodic memory retrieval.[2] Increased size of posterior ventral cingulate cortex is related to declines in working memory performance.[3] The posterior cingulate cortex has been strongly implicated as a key part of several intrinsic control networks.[4][5]
Anatomy
Location and boundaries
The posterior cingulate cortex is the backmost part of the cingulate cortex, lying behind the anterior cingulate cortex. The PCC forms a part of the posteromedial cortex, along with the retrosplenial cortex (Brodmann areas 29 and 30) and precuneus (located posterior and superior to the PCC). The PCC, together with the retrosplenial cortex, forms the retrosplenial gyrus. The posterior cingulate cortex is bordered by the following brain regions: the marginal ramus of the cingulate sulcus (superiorly), the corpus callosum (inferiorly), the parieto-occipital sulcus (posteriorly), and Brodmann area 24 (anteriorly).[4]
Cytoarchitectural organization
The posterior cingulate cortex is considered a paralimbic cortical structure, consisting of Brodmann areas 23 and 31. As part of the paralimbic cortex, it has fewer than six layers, placing its cell architecture in between the six-layered neocortex and the more primitive allocortex of core limbic structures. It has also been associated with the hippocampocentric subdivision of the paralimbic zone. The cytoarchitecture of the PCC is not entirely uniform, instead it contains distinct anterior and dorsal subregions, which are increasingly understood as distinct in function as well as cytoarchitectural structure.[4]
Structural connections
Nonhuman structure
In non-human primates the following structural connections of the posterior cingulate cortex are well documented:[4]
- Reciprocal connection with other regions of the posteromedial cortex.
- High connectivity to other paralimbic and limbic structures.
- Reciprocal connections to the medial temporal lobe.
- Dense connections to the hippocampal formation, the parahippocampal cortex, the ventromedial prefrontal cortex, and subgenual parts of the anterior cingulate cortex.
- Prominent connections to the areas of heteromodal association in the front, temporal, and parietal lobes.
- Strong reciprocal connections to the dorsolateral prefrontal cortex (roughly Brodmann area 46) and the frontal poles (Brodmann areas 10 and 11).
- Less prominent connections to Brodmann areas 9/46, 8, and 9.
- Connections to the dorsal parts of the anterior cingulate cortex.
- Dense connections the thalamus, in the form of a continuous strip crossing numerous pulvinar nuclei, and the striatum.
As is true in other areas of the posteromedial cortex, the posterior cingulate cortex has no apparent connections to primary sensory or motor areas. Thus, it is unlikely to be involved in low-level sensory or motor processing.[4]
Human structure
While many of the connections in non-human primates may be present in humans, they are less well documented. Studies have shown strong reciprocal connections to medial temporal lobe memory structures, such as the entorhinal cortex and the parahippocampal gyrus, which is known to be involved in associative learning and episodic memory.[6] The human posterior cingulate cortex is also connected to areas involved in emotion and social behavior, attention (the lateral intraparietal cortex and precuneus), and learning and motivation (the anterior and lateral thalamic nucleus, caudate nucleus, orbitofrontal cortex, and anterior cingulate cortex).[5][7]
Function
The posterior cingulate cortex is highly connected and one of the most metabolically active regions in the brain, but there is no consensus as to its cognitive role.[4][5] Cerebral blood flow and metabolic rate in the posterior cingulate cortex are approximately 40% higher than average across the brain. The posterior cingulate cortex also has high functional connectivity, signifying extensive intrinsic connectivity networks (networks of brain regions involved in a range of tasks that share common spatio-temporal patterns of activity).[4]
Emotion and memory
The posterior cingulate cortex has been linked by lesion studies to spatial memory, configural learning, and maintenance of discriminative avoidance learning.[6] More recently the PCC was shown to display intense activity when autobiographical memories (such as those concerning friends and family) are recalled successfully. In a study involving autobiographical recollection, the caudal part of the left posterior cingulate cortex was the only brain structure highly active in all subjects.[6] Furthermore, the PCC does not show this same activation during attempted but unsuccessful retrieval, implying an important role in successful memory retrieval (see below: Alzheimer’s Disease).[6]
The posterior cingulate cortex has also been firmly linked to emotional salience.[6][7] Thus, it has been hypothesized that the emotional importance of autobiographical memories may contribute to the strength and consistency of activity in the posterior cingulate cortex upon successful recollection of these memories.[6] The posterior cingulate cortex is significantly bilaterally activated by emotional stimuli, independent of valence (positive or negative). This is in contrast to other structures in the limbic system, such as the amygdala, which responded disproportionately to negative stimuli, or the left frontal pole, which activated only in response to positive stimuli. These results support the hypothesis that the posterior cingulate cortex mediates interactions between emotion and memory.
Intrinsic control networks
The posterior cingulate cortex exhibits connectivity with a wide range of intrinsic control networks. Its most widely known role is as a central node in the default mode network (DMN). The default mode network (and the PCC) is highly reactive and quickly deactivates during tasks with externally directed, or presently centered, attention (such as working memory or meditation).[4][8] Conversely, the DMN is active when attention is internally directed (during episodic memory retrieval, planning, and daydreaming). A failure of the DMN to deactivate at proper times is associated with poor cognitive function, thereby indicating its importance in attention.[4]
In addition to the default mode network, the posterior cingulate cortex is also involved in the dorsal attention network (a top-down control of visual attention and eye movement) and the frontoparietal control network (involved in executive motor control).[4] Furthermore, fMRI studies have shown that the posterior cingulate cortex activates during visual tasks when some form of monetary incentive is involved, essentially functioning as a neural interface between motivation-related areas and top-down control of visual attention.[9][10]
The relationship between these networks in and within the PCC is not clearly understood. When activity increases in the dorsal attention network and the frontoparietal control network, it must simultaneously decrease in the DMN in a closely correlated way. This anti-correlated pattern is indicative of the various differences and importance of subregions in the posterior cingulate cortex.[4]
Considering the PCC's relation with the DMN, since suppressed posterior cingulate activity favors low cognitive introspection and higher external attention while increased activity indicates memory retrieval and planning, it has been hypothesized that this brain region is heavily involved in noticing internal and external changes and facilitating novel behavior or thought in response. High activity, then, would indicate continued operation with the current cognitive set, while lower activity would indicate exploration, flexibility, and renewed learning.[5]
An alternative hypothesis is focused more on the difference between the dorsal and ventral subregions and takes into consideration their functional separation. In this model, the PCC is hypothesized to take a chief regulatory role in focusing internal and external attention. Mounting evidence that the posterior cingulate cortex is involved in both integrating memories of experiences and initiating a signal to change behavioral strategies supports this hypothesis. Under this model, the PCC plays a crucial role in controlling state of arousal, the breadth of focus, and the internal or external focus of attention. This hypothesis emphasizes the PCC as a dynamic network, rather than a fixed and unchanging structure.[4]
While both of the hypotheses are the result of scientific studies, the role of the PCC is still not well understood and there remains much work to be done to investigate the extent of their veracity.[4][5]
Meditation
From neuroimaging and subjective descriptions, the PCC has been found to be activated during self-related thinking and deactivated during meditation.[11] Using generative topographic mapping, it was further found that undistracted, effortless mind wandering corresponded with PCC activation, whereas distracted and controlled awareness corresponded with PCC deactivation.[11] These results track closely with findings about the PCC's role in the DMN.
Disorders
Structural and functional abnormalities in the PCC result in a range of neurological and psychiatric disorders. The PCC likely integrates and mediates information in the brain. Therefore, the functional abnormalities of the PCC might be an accumulation of remote and widespread damage in the brain.[4]
Alzheimer's disease
The PCC is commonly affected by neurodegenerative disease.[12] In fact, reduced metabolism in the PCC is an early sign of Alzheimer's disease, and is frequently present before a clinical diagnosis.[4] The reduced metabolism in the PCC is typically one part in a diffuse pattern of metabolic dysfunction in the brain that includes medial Temporal lobe structures and the anterior thalamus, and these metabolic abnormalities may be due to damage in isolated but connected regions.[4] For instance, Meguro et al. (1999) show that experimental damage of the rhinal cortex results in hypometabolism of the PCC.[13] In Alzheimer's disease, the metabolic abnormality is linked to Amyloid deposition and brain Atrophy with a spatial distribution that resembles the nodes of the Default mode network (DMN).[4] In early Alzheimer's, the functional connectivity within the DMN is reduced and affects the connection between the PCC and the hippocampus, and these altered patterns can reflect ApoE genetic status (a risk factor associated with the disease).[4] It has been found that neurodegenerative diseases spread 'prion-like' through the brain.[4] For example, when the proteins amyloid-b and TDP-43 are in their abnormal form, they spread trans-synaptically and are associated with Neurodegeneration.[4] This transmission of abnormal protein would be constrained by the organization of white matter connections and could potentially explain the spatial distribution of Alzheimer's pathology within the DMN.[4] In Alzheimer’s disease, the topology of white matter connectivity helps in predicting atrophic patterns,[14] which could explain why the PCC is affected in the early stages of the disease.[4]
Autism spectrum disorder
Autism spectrum disorders (ASDs) are associated with metabolic and functional abnormalities of the PCC. Individuals with ASDs show reduction in metabolism, exhibit abnormal functional responses and demonstrate reductions in functional connectivity.[4] One study showed these reductions are prominent in the PCC.[15] Studies have shown that the abnormalities in cingulate responses during interpersonal interaction correlate with the severity of autistic symptoms, and the failure to show task dependent deactivation in the PCC correlates with overall social function.[4] Finally, post-mortem studies show that the PCCs of patients with ASDs have cytoarchitectonic abnormalities, as well as reduced levels of GABA A receptors, and benzodiazepine binding sites[4]
Attention deficit hyperactivity disorder
It has been suggested that ADHD is a disorder of the DMN, where neural systems are disrupted by uncontrolled activity that leads to attentional lapses.[16] Nakao et al. (2011) performed a meta-analysis of structural MRI studies and found that patients with ADHD exhibit an increased left PCC,[17] which could mean that developmental abnormalities affect the PCC. In fact, PCC function is abnormal in ADHD.[4] Within the DMN, functional connectivity is reduced and the resting state activity is a way to diagnose children with ADHD.[4] Treatment for ADHD, includes psychostimulant medication that directly affects PCC activity.[4] Other studies looking at medications for abnormalities in the PCC found that the PCC may only respond to stimulant treatments and that the effectiveness of medication can be dependent on motivation levels.[4] Furthermore, ADHD has been associated with the gene SNAP25. In healthy children, SNAP25 polymorphism is linked to working memory capacity, altered PCC structure, and task-dependent PCC deactivation patterns on working memory task.[18]
Depression
Abnormal PCC functional connectivity has been linked to major depression. However, the conclusions of the studies vary. One found increased PCC functional connectivity,[19] while another showed that untreated patients had decreased functional connectivity from the PCC to the caudate.[20] Other studies have looked at interactions between the PCC and the sub-genual cingulate region (Brodmann area 25), a region of the brain that potentially causes depression.[4] The anterior node of the DMN is formed, in part, by the highly connected PCC and Brodmann area 25. The two regions are metabolically overactive in treatment resistant major depression.[21] The link between the activity in the PCC and Brodmann area 25 correlates with rumination, a feature of depression.[22] This link between the two regions could influence medication responses in patients. Already, it has been found that both regions show alterations in metabolism after antidepressant treatment. Furthermore, patients who undergo Deep brain stimulation, had increased glucose metabolism and cerebral flow in the PCC, while also altering Brodmann area 25.[4]
Schizophrenia
Abnormal activity in the PCC has been linked to schizophrenia, a mental disorder with common symptoms such as hallucinations, delusions, disorganized thinking, and a lack of emotional intelligence. What is common between symptoms is that they have to do with an inability to distinguish between internal and external events; they lack insight. Two PET studies on patients with schizophrenia showed abnormal metabolism in the PCC. One study found that glucose metabolism was decreased in schizophrenics,[23] while another found abnormal glucose metabolism that was highly correlated in the pulvinar and the PCC.[24] In the latter study, thalamic interactions with the frontal lobes were reduced, which could mean that schizophrenia affects thalamocortical connections. Further abnormalities in the PCC, abnormal NMDA, cannabinoid, and DABAergic receptor binding have been found with post-mortem autoradiography of schizophrenics.[25] Abnormalities in the structure and white matter connections of the PCC have also been found in schizophrenic patients. Schizophrenics with a poor outcome often have reduced PCC volume.[24] It has further been found that white matter abnormalities in the cingulum bundle, a structure that connects the PCC to other limbic structures.[26] In functional MRI studies, there is more evidence of abnormal PCC function. There can be increases and decreases in the functional connectivity.[27] There are also abnormal PCC responses during task performance.[28] All of the previously mentioned network abnormalities may be the reason for psychotic symptoms. Recently it was found that the psychedelic drug psilocybin induces an altered state of consciousness and is related to abnormal metabolism and functional connectivity of the PCC, as well as a reduction in the strength of anti-correlations between the DMN and the FPCN.[29] These networks contribute to internal and external cognition, so abnormalities in these networks might be responsible for the psychosis in schizophrenia.
Traumatic brain injury
After traumatic brain injury (TBI), abnormalities have been shown in the PCC. Often, head injuries produce widespread axonal injury that disconnect brain regions and lead to cognitive impairment. This is also related to reduced metabolism within the PCC.[30] Studies have looked at performance on simple choice reaction time tasks after TBIs.[31] This study, in particular, found that the pattern of functional connectivity from the PCC to the rest of the DMN could predict TBI impairments even before symptoms had manifested. They also found that greater damage to the cingulum bundle, that connects the PCC to the anterior DMN, was linked to impairments of sustained attention. In a subsequent study, it was found that TBIs are related to a difficulty in switching from automatic to controlled responses.[32] Within selected tasks, patients with TBIs showed impaired motor inhibition that was associated with failure to rapidly reactive the PCC. Collectively, this suggests that the failure to control the PCC/DMN activity can lead to attentional lapses in TBI patients.
See also
References
- ↑ R Leech; R Braga; DJ Sharp (2013). "Echoes of the brain within the posterior cingulate cortex". The Journal of Neuroscience. 32 (1): 215–222. doi:10.1523/JNEUROSCI.3689-11.2012.
- ↑ Nielsen FA, Balslev D, Hansen LK (2005). "Mining the posterior cingulate: segregation between memory and pain components". NeuroImage. 27 (3): 520–532. doi:10.1016/j.neuroimage.2005.04.034. PMID 15946864.
- ↑ Kozlovskiy SA, Vartanov AV, Nikonova EY, Pyasik MM, Velichkovsky BM (2012). "The Cingulate Cortex and Human Memory Processes". Psychology in Russia: State of the Art. 5: 231–243. doi:10.11621/pir.2012.0014.
- 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 Leech R, Sharp DJ (July 2013). "The role of the posterior cingulate cortex in cognition and disease". Brain. 137 (Pt 1): 12–32. doi:10.1093/brain/awt162. PMID 23869106.
- 1 2 3 4 5 Pearson, John M.; Heilbronner, Sarah R.; Barack, David L.; Hayden, Benjamin Y.; Platt, Michael L. (April 2011). "Posterior cingulate cortex: adapting behavior to a changing world". Trends in Cognitive Sciences. 15 (4): 143–151. doi:10.1016/j.tics.2011.02.002.
- 1 2 3 4 5 6 Maddock, R. J.; A. S. Garrett; M. H. Buonocore (2001). "Remembering Familiar People: The Posterior Cingulate Cortex and Autobiographical Memory Retrieval". Neuroscience. 104 (3): 667–676. doi:10.1016/s0306-4522(01)00108-7.
- 1 2 Maddock, Richard J.; Garrett, Amy S.; Buonocore, Michael H. (January 2003). "Posterior cingulate cortex activation by emotional words: fMRI evidence from a valence decision task". Human Brain Mapping. 18 (1): 30–41. doi:10.1002/hbm.10075.
- ↑ Brewer, Judson A.; Garrison, Kathleen A.; Whitfield-Gabrieli, Susan (2013). "What about the "Self" is Processed in the Posterior Cingulate Cortex?". Frontiers in Human Neuroscience. 7. doi:10.3389/fnhum.2013.00647.
- ↑ Small, Dana A.; Gitelman, Darren; Simmons, Katharine; Bloise, Suzanne; Parrish, Todd; Mesulam, Marsel M. (2005). "Monetary Incentives Enhance Processing in Brain Regions Mediating Top-down Control of Attention". Cerebral Cortex. 15: 1855–1865. doi:10.1093/cercor/bhi063.
- ↑ Engelmann, Jan B.; Damaraju, Eswar; Padmala, Srikanth; Pessoa, Luiz (2009). "Combined Effects of Attention and Motivation on Visual Task Performance: Transient and Sustained Motivational Effects". Frontiers in Human Neuroscience. 3. doi:10.3389/neuro.09.004.2009.
- 1 2 Garrison, KA; Santoyo, JF; Davis, JH; Thornhill TA, 4th; Kerr, CE; Brewer, JA (2013). "Effortless awareness: using real time neurofeedback to investigate correlates of posterior cingulate cortex activity in meditators' self-report.". Frontiers in Human Neuroscience. 7: 440. doi:10.3389/fnhum.2013.00440. PMID 23964222. Cite uses deprecated parameter
|coauthors=(help) - ↑ Buckner, RL; Andrews-Hanna JR; Schacter DL (2008). "The brain's default network: anatomy, function, and relevance to disease". Ann N Y Acad Sci. 1124: 1–38. doi:10.1196/annals.1440.011. PMID 18400922.
- ↑ Meguro, K; Blaizot X, Kondoh Y, Le Mestric C, Baron JC, Chavoix C. (1999). "Neocortical and hippocampal glucose hypometabolism following neurotoxic lesions of the entorhinal and perirhinal cortices in the non-human primate as shown by PET. Implications for Alzheimer's disease.". Brain. 122 (Pt 8): 1519–31. doi:10.1093/brain/122.8.1519. Cite uses deprecated parameter
|coauthors=(help) - ↑ Raj, A; Kuceyeski A; Weiner M (2012). "A network diffusion model of disease progression in dementia". Neuron. 73: 1204–15. doi:10.1016/j.neuron.2011.12.040.
- ↑ Cherkassky, VL; Kana, RK; Keller, TA; Just, MA (Nov 6, 2006). "Functional connectivity in a baseline resting-state network in autism.". NeuroReport. 17 (16): 1687–90. doi:10.1097/01.wnr.0000239956.45448.4c. PMID 17047454.
- ↑ Sonuga-Barke, EJ; Castellanos, FX (2007). "Spontaneous attentional fluctuations in impaired states and pathological conditions: a neurobiological hypothesis.". Neuroscience and biobehavioral reviews. 31 (7): 977–86. doi:10.1016/j.neubiorev.2007.02.005. PMID 17445893.
- ↑ Nakao, T; Radua, J; Rubia, K; Mataix-Cols, D (November 2011). "Gray matter volume abnormalities in ADHD: voxel-based meta-analysis exploring the effects of age and stimulant medication.". The American Journal of Psychiatry. 168 (11): 1154–63. doi:10.1176/appi.ajp.2011.11020281. PMID 21865529.
- ↑ Latasch, L; Christ, R (March 1988). "[Problems in anesthesia of drug addicts].". Der Anaesthesist. 37 (3): 123–39. PMID 3289412.
- ↑ Zhou, Y; Yu, C; Zheng, H; Liu, Y; Song, M; Qin, W; Li, K; Jiang, T (March 2010). "Increased neural resources recruitment in the intrinsic organization in major depression.". Journal of Affective Disorders. 121 (3): 220–30. doi:10.1016/j.jad.2009.05.029. PMID 19541369.
- ↑ Bluhm, R; Williamson, P; Lanius, R; Théberge, J; Densmore, M; Bartha, R; Neufeld, R; Osuch, E (December 2009). "Resting state default-mode network connectivity in early depression using a seed region-of-interest analysis: decreased connectivity with caudate nucleus.". Psychiatry and clinical neurosciences. 63 (6): 754–61. doi:10.1111/j.1440-1819.2009.02030.x. PMID 20021629.
- ↑ Mayberg, HS; Liotti, M; Brannan, SK; McGinnis, S; Mahurin, RK; Jerabek, PA; Silva, JA; Tekell, JL; Martin, CC; Lancaster, JL; Fox, PT (May 1999). "Reciprocal limbic-cortical function and negative mood: converging PET findings in depression and normal sadness.". The American Journal of Psychiatry. 156 (5): 675–82. PMID 10327898.
- ↑ Berman, MG; Peltier, S; Nee, DE; Kross, E; Deldin, PJ; Jonides, J (October 2011). "Depression, rumination and the default network.". Social cognitive and affective neuroscience. 6 (5): 548–55. doi:10.1093/scan/nsq080. PMID 20855296.
- ↑ Haznedar, MM; Buchsbaum, MS; Hazlett, EA; Shihabuddin, L; New, A; Siever, LJ (Dec 1, 2004). "Cingulate gyrus volume and metabolism in the schizophrenia spectrum.". Schizophrenia Research. 71 (2-3): 249–62. doi:10.1016/j.schres.2004.02.025. PMID 15474896.
- 1 2 Mitelman, SA; Byne, W; Kemether, EM; Hazlett, EA; Buchsbaum, MS (September 2005). "Metabolic disconnection between the mediodorsal nucleus of the thalamus and cortical Brodmann's areas of the left hemisphere in schizophrenia.". The American Journal of Psychiatry. 162 (9): 1733–5. doi:10.1176/appi.ajp.162.9.1733. PMID 16135634.
- ↑ Newell, KA; Zavitsanou, K; Huang, XF (Aug 22, 2005). "Ionotropic glutamate receptor binding in the posterior cingulate cortex in schizophrenia patients.". NeuroReport. 16 (12): 1363–7. doi:10.1097/01.wnr.0000174056.11403.71. PMID 16056140.
- ↑ Kubicki, M; McCarley, RW; Nestor, PG; Huh, T; Kikinis, R; Shenton, ME; Wible, CG (December 2003). "An fMRI study of semantic processing in men with schizophrenia.". NeuroImage. 20 (4): 1923–33. doi:10.1016/s1053-8119(03)00383-5. PMID 14683698.
- ↑ Liang, M; Zhou, Y; Jiang, T; Liu, Z; Tian, L; Liu, H; Hao, Y (Feb 6, 2006). "Widespread functional disconnectivity in schizophrenia with resting-state functional magnetic resonance imaging.". NeuroReport. 17 (2): 209–13. doi:10.1097/01.wnr.0000198434.06518.b8. PMID 16407773.
- ↑ Whitfield-Gabrieli, S; Thermenos, HW; Milanovic, S; Tsuang, MT; Faraone, SV; McCarley, RW; Shenton, ME; Green, AI; Nieto-Castanon, A; LaViolette, P; Wojcik, J; Gabrieli, JD; Seidman, LJ (Jan 27, 2009). "Hyperactivity and hyperconnectivity of the default network in schizophrenia and in first-degree relatives of persons with schizophrenia.". Proceedings of the National Academy of Sciences of the United States of America. 106 (4): 1279–84. doi:10.1073/pnas.0809141106. PMC 2633557
 . PMID 19164577.
. PMID 19164577. - ↑ Carhart-Harris, RL; Erritzoe, D; Williams, T; Stone, JM; Reed, LJ; Colasanti, A; Tyacke, RJ; Leech, R; Malizia, AL; Murphy, K; Hobden, P; Evans, J; Feilding, A; Wise, RG; Nutt, DJ (Feb 7, 2012). "Neural correlates of the psychedelic state as determined by fMRI studies with psilocybin.". Proceedings of the National Academy of Sciences of the United States of America. 109 (6): 2138–43. doi:10.1073/pnas.1119598109. PMC 3277566. PMID 22308440. Cite uses deprecated parameter
|coauthors=(help) - ↑ Nakashima, T; Nakayama, N; Miwa, K; Okumura, A; Soeda, A; Iwama, T (February 2007). "Focal brain glucose hypometabolism in patients with neuropsychologic deficits after diffuse axonal injury.". AJNR. American journal of neuroradiology. 28 (2): 236–42. PMID 17296986.
- ↑ Bonnelle, V; Leech, R; Kinnunen, KM; Ham, TE; Beckmann, CF; De Boissezon, X; Greenwood, RJ; Sharp, DJ (Sep 21, 2011). "Default mode network connectivity predicts sustained attention deficits after traumatic brain injury.". The Journal of neuroscience : the official journal of the Society for Neuroscience. 31 (38): 13442–51. doi:10.1523/JNEUROSCI.1163-11.2011. PMID 21940437. Cite uses deprecated parameter
|coauthors=(help) - ↑ Bonnelle, V; Ham, TE; Leech, R; Kinnunen, KM; Mehta, MA; Greenwood, RJ; Sharp, DJ (Mar 20, 2012). "Salience network integrity predicts default mode network function after traumatic brain injury.". Proceedings of the National Academy of Sciences of the United States of America. 109 (12): 4690–5. doi:10.1073/pnas.1113455109. PMID 22393019.
External links
| Wikimedia Commons has media related to Posterior cingulate cortex. |
- Posterior cingulate in the Brede Database at the Technical University of Denmark
For details regarding MRI definitions of the cingulate cortex based on the Desikan-Killiany Brain atlas, see:
- Desikan RS, Ségonne F, Fischl B, Quinn BT, Dickerson BC, Blacker D, Buckner RL, Dale AM, Maguire RP, et al. (Jul 2006). "An automated labeling system for subdividing the human cerebral cortex on MRI scans into gyral based regions of interest". NeuroImage. 31 (3): 968–80. doi:10.1016/j.neuroimage.2006.01.021.