Nitrogen fixation
Nitrogen fixation is a process in which nitrogen (N2) in the Earth's atmosphere is converted into ammonia (NH3).[1] Atmospheric nitrogen or molecular dinitrogen (N2) is relatively inert: it does not easily react with other chemicals to form new compounds. The fixation process frees nitrogen atoms from their triply bonded diatomic form, N≡N, to be used in other ways.
Nitrogen fixation, natural and synthetic, is essential for all forms of life because nitrogen is required to biosynthesize basic building blocks of plants, animals and other life forms, e.g., nucleotides for DNA and RNA, the coenzyme nicotinamide adenine dinucleotide for its role in metabolism (transferring electrons between molecules), and amino acids for proteins. Therefore, as part of the nitrogen cycle, it is essential for agriculture and the manufacture of fertilizer. It is also, indirectly, relevant to the manufacture of all chemical compounds that contain nitrogen, which includes explosives, most pharmaceuticals, dyes, etc. Nitrogen fixation occurs naturally in the soil by nitrogen fixing bacteria (for example, Azotobacter affiliated with some plants, especially legumes). Some nitrogen-fixing bacteria have very close relationships with plants, referred to as symbiotic nitrogen fixation. Looser relationships between nitrogen-fixing bacteria and plants are often referred to as associative or non-symbiotic, as seen in nitrogen fixation occurring on rice roots. It also occurs naturally in the air by means of lightning.[2][3]
All biological nitrogen fixation is done by way of metalloenzymes called nitrogenase. These enzymes contain iron, often with a second metal, usually molybdenum but sometimes vanadium. Microorganisms that can fix nitrogen are prokaryotes (both bacteria and archaea, distributed throughout their respective kingdoms) called diazotrophs. Some higher plants, and some animals (termites), have formed associations (symbiosis) with diazotrophs.
Biological nitrogen fixation
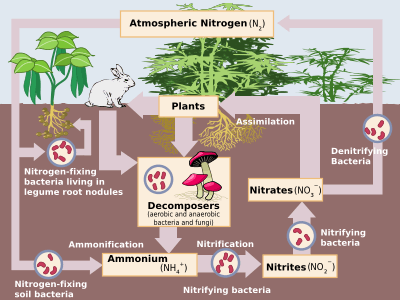
Biological nitrogen fixation was discovered by the German agronomist Hermann Hellriegel[4] and Dutch microbiologist Martinus Beijerinck.[5] Biological nitrogen fixation (BNF) occurs when atmospheric nitrogen is converted to ammonia by an enzyme called a nitrogenase.[1] The overall reaction for BNF is:
- N2 + 8 H+ + 8 e− → 2 NH3 + H2
The process is coupled to the hydrolysis of 16 equivalents of ATP and is accompanied by the co-formation of one molecule of H2.[6] The conversion of N2 into ammonia occurs at a cluster called FeMoco, an abbreviation for the iron-molybdenum cofactor. The mechanism proceeds via a series of protonation and reduction steps wherein the FeMoco active site hydrogenates the N2 substrate.[7]
In free-living diazotrophs, the nitrogenase-generated ammonium is assimilated into glutamate through the glutamine synthetase/glutamate synthase pathway.
The microbial genes required for nitrogen fixation are widely distributed in diverse environments.[8][9]
Enzymes responsible for nitrogenase action are very susceptible to destruction by oxygen. For this reason, many bacteria cease production of the enzyme in the presence of oxygen. Many nitrogen-fixing organisms exist only in anaerobic conditions, respiring to draw down oxygen levels, or binding the oxygen with a protein such as leghemoglobin.[1]
Microorganisms that fix nitrogen
Diazotrophs are a diverse group of prokaryotes that includes cyanobacteria (e.g. the highly significant Trichodesmium and Cyanothece), green sulfur bacteria, along with Azotobacteraceae, rhizobia and Frankia.
Cyanobacteria inhabit nearly all illuminated environments on Earth and play key roles in the carbon and nitrogen cycle of the biosphere. In general, cyanobacteria are able to utilize a variety of inorganic and organic sources of combined nitrogen, like nitrate, nitrite, ammonium, urea, or some amino acids. Several cyanobacterial strains are also capable of diazotrophic growth, an ability that may have been present in their last common ancestor in the Archean eon.[10] Nitrogen fixation by cyanobacteria in coral reefs can fix twice the amount of nitrogen as on land—around 1.8 kg of nitrogen is fixed per hectare per day (around 660 kg/ha/year). The colonial marine cyanobacterium Trichodesmium is thought to fix nitrogen on such a scale that it accounts for almost half of the nitrogen fixation in marine systems on a global scale.[11]
Root nodule symbioses
Legume family
Plants that contribute to nitrogen fixation include the legume family – Fabaceae – with taxa such as kudzu, clovers, soybeans, alfalfa, lupines, peanuts, and rooibos. They contain symbiotic bacteria called rhizobia within nodules in their root systems, producing nitrogen compounds that help the plant to grow and compete with other plants. When the plant dies, the fixed nitrogen is released, making it available to other plants; this helps to fertilize the soil.[1][12] The great majority of legumes have this association, but a few genera (e.g., Styphnolobium) do not. In many traditional and organic farming practices, fields are rotated through various types of crops, which usually include one consisting mainly or entirely of clover or buckwheat (non-legume family Polygonaceae), often referred to as "green manure".[13]
The efficiency of nitrogen fixation in soil is dependent on many factors, including the legume as well as air and and soil conditions. For example, nitrogen fixation by red clover can range from 50 - 200lb/acre depending on these variables.[14]
Inga alley farming relies on the leguminous genus Inga, a small tropical, tough-leaved, nitrogen-fixing tree.[15]
Non-leguminous

Although by far the majority of plants able to form nitrogen-fixing root nodules are in the legume family Fabaceae, there are a few exceptions:
- Parasponia, a tropical genus in the Cannabaceae also able to interact with rhizobia and form nitrogen-fixing nodules[16]
- Actinorhizal plants such as alder and bayberry can also form nitrogen-fixing nodules, thanks to a symbiotic association with Frankia bacteria. These plants belong to 25 genera[17] distributed among 8 plant families.
The ability to fix nitrogen is far from universally present in these families. For instance, of 122 genera in the Rosaceae, only 4 genera are capable of fixing nitrogen. All these families belong to the orders Cucurbitales, Fagales, and Rosales, which together with the Fabales form a clade of eurosids. In this clade, Fabales were the first lineage to branch off; thus, the ability to fix nitrogen may be plesiomorphic and subsequently lost in most descendants of the original nitrogen-fixing plant; however, it may be that the basic genetic and physiological requirements were present in an incipient state in the last common ancestors of all these plants, but only evolved to full function in some of them:
| Family: Genera
Betulaceae: Alnus (alders) |
…… |
|
…… |
…… |
…… |
|
There are also several nitrogen-fixing symbiotic associations that involve cyanobacteria (such as Nostoc):
Endosymbiosis in diatoms
Rhopalodia gibba, a diatom algae, is a eukaryote with cyanobacterial N2-fixing endosymbiont organelles. The spheroid bodies reside in the cytoplasm of the diatoms and are inseparable from their hosts.[18][19]
Industrial nitrogen fixation
The possibility that atmospheric nitrogen reacts with certain chemicals was first observed by Desfosses in 1828. He observed that mixtures of alkali metal oxides and carbon react at high temperatures with nitrogen. With the use of barium carbonate as starting material the first commercially used process became available in the 1860s developed by Margueritte and Sourdeval. The resulting barium cyanide could be reacted with steam yielding ammonia. In 1898 Adolph Frank and Nikodem Caro decoupled the process and first produced calcium carbide and in a subsequent step reacted it with nitrogen to calcium cyanamide. The Ostwald process for the production of nitric acid was discovered in 1902. Frank-Caro process and Ostwald process dominated the industrial fixation of nitrogen until the discovery of the Haber process in 1909.[20][21] Prior to 1900, Nikola Tesla also experimented with the industrial fixation of nitrogen "by using currents of extremely high frequency or rate of vibration".[22][23]
Haber process

Artificial fertilizer production is now the largest source of human-produced fixed nitrogen in the Earth's ecosystem. Ammonia is a required precursor to fertilizers, explosives, and other products. The most common method is the Haber process. The Haber process requires high pressures (around 200 atm) and high temperatures (at least 400 °C), routine conditions for industrial catalysis. This highly efficient process uses natural gas as a hydrogen source and air as a nitrogen source.[24]
Much research has been conducted on the discovery of catalysts for nitrogen fixation, often with the goal of reducing the energy required for this conversion. However, such research has thus far failed to even approach the efficiency and ease of the Haber process. Many compounds react with atmospheric nitrogen to give dinitrogen complexes. The first dinitrogen complex to be reported was Ru(NH3)5(N2)2+.[25]
Ambient nitrogen reduction
Catalytic chemical nitrogen fixation at ambient conditions is an ongoing scientific endeavor. Guided by the example of nitrogenase, this area of homogeneous catalysis is ongoing, with particular emphasis on hydrogenation to give ammonia.[26]
Metallic lithium has long been known for burning in an atmosphere of nitrogen and then converting to lithium nitride. Hydrolysis of the resulting nitride gives ammonia. In a related process, trimethylsilyl chloride, lithium, and nitrogen react in the presence of a catalyst to give tris(trimethylsilyl)amine. Tris(trimethylsilyl)amine can then be used for reaction with α,δ,ω-triketones to give tricyclic pyrroles.[27] Processes involving lithium metal are however of no practical interest since they are noncatalytic and re-reducing the Li+ ion residue is difficult.
Beginning in the 1960s several homogeneous systems were identified that convert nitrogen to ammonia, sometimes even catalytically but often operating via ill-defined mechanisms. The original discovery is described in an early review:
"Vol'pin and co-workers, using a non-protic Lewis acid, aluminium tribromide, were able to demonstrate the truly catalytic effect of titanium by treating dinitrogen with a mixture of titanium tetrachloride, metallic aluminium, and aluminium tribromide at 50 °C, either in the absence or in the presence of a solvent, e.g. benzene. As much as 200 mol of ammonia per mol of TiCl4 was obtained after hydrolysis.…"[28]
The quest for well defined intermediates led to the characterization of many transition metal dinitrogen complexes. Few of these well defined complexes function catalytically, their behavior illuminated likely stages in nitrogen fixation. Most fruitful of all of these early studies focused on M(N2)2(dppe)2 (M = Mo, W). For example, double protonation of such low valent complexes gave intermediates with the linkage M=N−NH2. In 1995, a molybdenum(III) amido complex was discovered that cleaved N2 to give the corresponding molybdenum(VI) nitride.[29] This and related terminal nitrido complexes have been used to make nitriles.[30]

In 2003 a related molybdenum amido complex was found to catalyze the reduction of N2. In addition to a source of protons, the catalyst requires a strong reducing agent.[31][32][33][34] However, this catalytic reduction fixates only a few nitrogen molecules. In these systems, like the biological one, hydrogen is provided to the substrate heterolytically, by means of protons and reducing equivalents rather than with H2 itself.
In 2011 Arashiba et al. reported yet another system with a catalyst again based on molybdenum but with a diphosphorus pincer ligand.[35] Photolytic nitrogen splitting is also considered.[36][37][38][39][40]
See also
- Birkeland–Eyde process: an industrial fertiliser production process
- Denitrification: an organic process of nitrogen release
- George Washington Carver: an American botanist
- Nif gene: a gene found in nitrogen fixing bacteria
- Nitrification: biological production of nitrogen
- Nitrogen cycle: the flow and transformation of nitrogen through the environment
- Nitrogen deficiency
- Nitrogen fixation package for quantitative measurement of nitrogen fixation by plants
- Nitrogenase: enzymes used by organisms to fix nitrogen
- Ostwald process: a chemical process for making nitric acid HNO3
- Push–pull technology: the use of both repellent and attractive organisms in agriculture
References
- 1 2 3 4 Postgate, J. (1998). Nitrogen Fixation (3rd ed.). Cambridge: Cambridge University Press.
- ↑ Slosson, Edwin (1919). Creative Chemistry. New York, NY: The Century Co. pp. 19–37.
- ↑ Hill, R. D.; Rinker, R. G.; Wilson, H. Dale (1979). "Atmospheric Nitrogen Fixation by Lightning". J. Atmos. Sci. 37 (1): 179–192. doi:10.1175/1520-0469(1980)037<0179:ANFBL>2.0.CO;2.
- ↑ Hellriegel, H.; Wilfarth, H. (1888). Untersuchungen über die Stickstoffnahrung der Gramineen und Leguminosen [Studies on the nitrogen intake of Gramineae and Leguminosae]. Berlin: Buchdruckerei der "Post" Kayssler & Co.
- ↑ Beijerinck, M. W. (1901). "Über oligonitrophile Mikroben" [On oligonitrophilic microbes]. Centralblatt für Bakteriologie, Parasitenkunde, Infektionskrankheiten und Hygiene. 7 (2): 561–582.
- ↑ Chi Chung, Lee; Markus W., Ribbe; Yilin, Hu (2014). "Chapter 7. Cleaving the N,N Triple Bond: The Transformation of Dinitrogen to Ammonia by Nitrogenases". In Kroneck, Peter M. H.; Sosa Torres, Martha E. The Metal-Driven Biogeochemistry of Gaseous Compounds in the Environment. Metal Ions in Life Sciences. 14. Springer. pp. 147–174. doi:10.1007/978-94-017-9269-1_6.
- ↑ Hoffman, B. M.; Lukoyanov, D.; Dean, D. R.; Seefeldt, L. C. (2013). "Nitrogenase: A Draft Mechanism". Acc. Chem. Res. 46: 587–595. doi:10.1021/ar300267m.
- ↑ Gaby, J. C.; Buckley, D. H. (2011). "A global census of nitrogenase diversity". Environ. Microbiol. 13 (7): 1790–1799. doi:10.1111/j.1462-2920.2011.02488.x. PMID 21535343.
- ↑ Hoppe, B.; Kahl, T.; Karasch, P.; Wubet, T.; Bauhus, J.; Buscot, F.; Krüger, D. (2014). "Network analysis reveals ecological links between N-fixing bacteria and wood-decaying fungi". PLoS ONE. 9 (2): e88141. Bibcode:2014PLoSO...988141H. doi:10.1371/journal.pone.0088141. PMC 3914916
 . PMID 24505405.
. PMID 24505405. - ↑ Latysheva, N.; Junker, V. L.; Palmer, W. J.; Codd, G. A.; Barker, D. (2012). "The evolution of nitrogen fixation in cyanobacteria". Bioinformatics. 28 (5): 603–606. doi:10.1093/bioinformatics/bts008.
- ↑ Bergman, B.; Sandh, G.; Lin, S.; Larsson, H.; Carpenter, E. J. (2012). "Trichodesmium – a widespread marine cyanobacterium with unusual nitrogen fixation properties". FEMS Microbiol. Rev. 37 (3): 1–17. doi:10.1111/j.1574-6976.2012.00352.x.
- ↑ Smil, Vaclav (2000). Cycles of Life. Scientific American Library.
- ↑ By who?
- ↑ "Nitrogen Fixation and Inoculation of Forage Legumes" (PDF).
- ↑ Elkan, Daniel (21 April 2004). "Slash-and-burn farming has become a major threat to the world's rainforest". The Guardian.
- ↑ Op den Camp, Rik; Streng, A.; De Mita, S.; Cao, Q.; Polone, E.; Liu, W.; Ammiraju, J. S. S.; Kudrna, D.; Wing, R.; Untergasser, A.; Bisseling, T.; Geurts, R. (2010). "LysM-Type Mycorrhizal Receptor Recruited for Rhizobium Symbiosis in Nonlegume Parasponia". Science. 331 (6019): 909–912. Bibcode:2011Sci...331..909O. doi:10.1126/science.1198181.
- ↑ Dawson, J. O. (2008). "Ecology of actinorhizal plants". Nitrogen-fixing Actinorhizal Symbioses. 6. Springer. pp. 199–234. doi:10.1007/978-1-4020-3547-0_8.
- ↑ Prechtl, Julia; Kneip, Christoph; Lockhart, Peter; Wenderoth, Klaus; Maier, Uwe-G. (2004). "Intracellular spheroid bodies of Rhopalodia gibba have nitrogen-fixing apparatus of cyanobacterial origin". Mol. Biol. Evol. 21 (8): 1477–1481. doi:10.1093/molbev/msh086.
- ↑ Nakayama, Takuro; Inagaki, Yuji (2014). "Unique genome evolution in an intracellular N2-fixing symbiont of a rhopalodiacean diatom". Acta Soc. Bot. Polon. 83 (4): 409–413. doi:10.5586/asbp.2014.046.
- ↑ Heinrich, H.; Nevbner, Rolf (1934). "Die Umwandlungsgleichung Ba(CN)2 → BaCN2 + C im Temperaturgebiet von 500 bis 1000 °C" [The conversion reaction Ba(CN)2 → BaCN2 + C in the temperature range from 500 to 1,000 °C]. Z. Elektrochem. Angew. Phys. Chem. 40 (10): 693–698. Retrieved 8 August 2016.
- ↑ Curtis, Harry Alfred (1932). Fixed nitrogen.
- ↑ ""The Problem of Increasing Human Energy" by Nikola Tesla". Retrieved 22 June 2016.
- ↑ Tesla, Nikola (1900). "The Problem of Increasing Human Energy". The Century Magazine. 60 (n.s. v. 38) (1900 May–Oct): 175.
- ↑ Vitousek, Peter M.; Aber, John; Howarth, Robert W.; Likens, Gene E.; Matson, Pamela A.; Schindler, David W.; Schlesinger, William H.; Tilman, G. David. "Human Alteration of the Global Nitrogen Cycle: Causes and Consequences" (PDF). US Environmental Protection Agency.
- ↑ Allen, A. D.; Senoff, C. V. (1965). "Nitrogenopentammineruthenium(II) complexes". J. Chem. Soc., Chem. Comm. (24): 621. doi:10.1039/C19650000621.
- ↑ Schrock, Richard R. (2006). "Reduction of dinitrogen". PNAS. 103 (46): 17087. doi:10.1073/pnas.0603633103.
- ↑ Brook, Michael A. (2000). Silicon in Organic, Organometallic, and Polymer Chemistry. New York: John Wiley & Sons, Inc. pp. 193–194.
- ↑ Chatt, J.; Leigh, G. J. (1972). "Nitrogen Fixation". Chem. Soc. Rev. 1: 121. doi:10.1039/cs9720100121.
- ↑ Laplaza, Catalina E.; Cummins, Christopher C. (1995). "Dinitrogen Cleavage by a Three-Coordinate Molybdenum(III) Complex". Science. 268 (5212): 861–863. doi:10.1126/science.268.5212.861.
- ↑ Curley, John J.; Sceats, Emma L.; Cummins, Christopher C. (2006). "A Cycle for Organic Nitrile Synthesis via Dinitrogen Cleavage". J. Am. Chem. Soc. 128 (43): 14036–14037. doi:10.1021/ja066090a.
- 1 2 Yandulov, Dmitry V.; Schrock, Richard R.; Rheingold, Arnold L.; Ceccarelli, Christopher; Davis, William M. (2003). "Synthesis and Reactions of Molybdenum Triamidoamine Complexes Containing Hexaisopropylterphenyl Substituents". Inorg. Chem. 42 (3): 796–813. doi:10.1021/ic020505l.
- ↑ Yandulov, Dmitry V.; Schrock, Richard R. (2003). "Catalytic Reduction of Dinitrogen to Ammonia at a Single Molybdenum Center". Science. 301 (5629): 76–78. doi:10.1126/science.1085326.
- ↑ The catalyst is derived from molybdenum(V) chloride and tris(2-aminoethyl)amine N-substituted with three very bulky hexa-isopropylterphenyl (HIPT) groups. Nitrogen adds end-on to the molybdenum atom, and the bulky HIPT substituents prevent the formation of the stable and nonreactive Mo−N=N−Mo dimer. In this isolated pocket is the Mo−N2. The proton donor is a pyridinium salt of weakly coordinating counter anion. The reducing agent is decamethylchromocene. All ammonia formed is collected as the HCl salt by trapping the distillate with a HCl solution.
- ↑ Note also that, although the dinitrogen complex is shown in brackets, this species can be isolated and characterized. Here the brackets do not indicate that the intermediate is not observed.
- ↑ Arashiba, Kazuya; Miyake, Yoshihiro; Nishibayashi, Yoshiaki (2011). "A molybdenum complex bearing PNP-type pincer ligands leads to the catalytic reduction of dinitrogen into ammonia". Nature Chem. 3: 120–125. doi:10.1038/nchem.906.
- ↑ Rebreyend, C.; de Bruin, B. (2014). "Photolytic N2 Splitting: A Road to Sustainable NH3 Production?". Angew. Chem. Int. Ed. doi:10.1002/anie.201409727.
- ↑ Solari, E.; Da Silva, C.; Iacono, B.; Hesschenbrouck, J.; Rizzoli, C.; Scopelliti, R.; Floriani, C. (2001). "Photochemical Activation of the N≡N Bond in a Dimolybdenum–Dinitrogen Complex: Formation of a Molybdenum Nitride". Angew. Chem. Int. Ed. 40: 3907–3909. doi:10.1002/1521-3773(20011015)40:20<3907::AID-ANIE3907>3.0.CO;2-#.
- ↑ Huss, Adam S.; Curley, John J.; Cummins, Christopher C.; Blank, David A. (2013). "Relaxation and Dissociation Following Photoexcitation of the (μ-N2)[Mo(N[t-Bu]Ar)3]2 Dinitrogen Cleavage Intermediate". J. Phys. Chem. B. 117 (5): 1429–1436. doi:10.1021/jp310122x.
- ↑ Kunkely, H.; Vogler, A. (2010). "Photolysis of Aqueous [(NH3)5Os(μ-N2)Os(NH3)5]5+: Cleavage of Dinitrogen by an Intramolecular Photoredox Reaction". Angew. Chem. Int. Ed. 49: 1591–1593. doi:10.1002/anie.200905026.
- ↑ Miyazaki, T.; Tanaka, H.; Tanabe, Y.; Yuki, M.; Nakajima, K.; Yoshizawa, K.; Nishibayashi, Y. (2014). "Cleavage and Formation of Molecular Dinitrogen in a Single System Assisted by Molybdenum Complexes Bearing Ferrocenyldiphosphine". Angew. Chem. Int. Ed. 53: 11488–11492. doi:10.1002/anie.201405673.
External links
- Hirsch, Ann M. (2009). "A Brief History of the Discovery of Nitrogen-fixing Organisms" (PDF). University of California, Los Angeles.
- "Marine Nitrogen Fixation laboratory". University of Southern California.