CFU-E
CFU-E is a hematopoietic colony forming unit. It arises from CFU-GEMM (via BFU-E,[1] which stands for "erythroid burst-forming units"[2]) and gives rise to proerythroblasts.
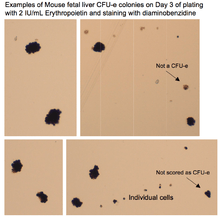
Understanding the murine CFU-e assay (analogous to human assay):
CFU-e is a stage of erythroid development between the BFU-e stage and the pro-erythroblast stage. CFU-e colony assay is designed to detect how many colony-forming-units of erythroid lineage there are in a hematopoietic tissue (bone marrow, spleen, or fetal liver), which may be reflective of the organism’s demand for oxygen delivery to the tissues or a hematopoietic disorder.
Early erythroid progenitors are found at a quite low frequency relative to later stages of erythroid differentiation, such as the pro-erythroblast and the basophilic erythroblast stages which can be detected by flowcytometry directly ex-vivo (Socolovsky et al. 2001 PMID 11719363). Furthermore, unlike for the pro-erythroblast and later stages of erythroid development, no truly reliable and unique positive flow-cytometric markers exist, though it is possible to use negative exclusion markers to deplete a cell population of other precursors and differentiated cells by cell sorting, thus greatly enriching it for the CFU-e activity (Prospective isolation and global gene expression analysis of the erythrocyte colony-forming unit, Terszowski G. et al. Blood 2005). CFU-e cells express Epo receptor, c-Kit (Stem cell factor receptor), transferrin receptor (CD71+), and are Ter119(glycophorin-A associated antigen)-negative. For the above reasons, the CFU-e assay, as inefficient and variable as it can often be, is still in use today.
Cells at the CFU-e stage express some erythropoietin receptor (EpoR), and thus can be induced to terminally differentiate in vitro in 2–3 days in the presence of only erythropoietin (Epo) (together with the basic contents of culture media: FBS, BSA in IMDM). Methylcellulose is a semisolid media additive that allows an investigator to stain (with diaminobenzidine reagent for hemoglobin) and then count individual colonies, each arising from a single plated progenitor that is at the CFU-e stage. By day 2 from the time of plating, each CFU-e colony will contain between 8 (minimum) and 64 hemoglobinized cells most of which are in their end-stage of erythroid differentiation. It is possible to see a small spectrum of hemoglobinization level and possibly cell size, indicating that some cells in the colony have achieved the end-stage faster than others.
Cell number in a colony is important because pro-erythroblast stage is also Epo-responsive (expresses Epo receptor), but the proliferative capacity of these cells is not as high, thus yielding a colony with fewer than 8 cells. Likewise, an earlier stage of erythroid differentiation may also yield colonies in Epo-only medium, but these colonies would likely be smaller and/or not hemoglobinized, since the stages before the CFU-e stage (MEP and BFU-e) require other factors (IL-3 etc.) and more time for growth that will also delay the terminal differentiation and hemoglobinization.
References
- ↑ Wu H, Liu X, Jaenisch R, Lodish HF (October 1995). "Generation of committed erythroid BFU-E and CFU-E progenitors does not require erythropoietin or the erythropoietin receptor". Cell. 83 (1): 59–67. doi:10.1016/0092-8674(95)90234-1. PMID 7553874.
- ↑ Marley SB, Lewis JL, Goldman JM, Gordon MY (June 1996). "Abnormal kinetics of colony formation by erythroid burst-forming units (BFU-E) in chronic myeloid leukaemia". Br. J. Haematol. 93 (4): 878–83. doi:10.1046/j.1365-2141.1996.d01-1738.x. PMID 8703820.
External links
- CFU-E at the US National Library of Medicine Medical Subject Headings (MeSH)